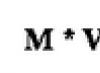Тесты
620-1. Скопление какой группы растений способствует заболачиванию почвы?
А) плауновидных
Б) хвощевидных
В) моховидных
Г) папортниковидных
Ответ
620-2. Стебель с листьями в процессе эволюции впервые появился у
А) водорослей
Б) моховидных
В) хвощевидных
Г) папоротниковидных
Ответ
620-3. Мхи представляют тупиковую ветвь в эволюции растений, так как
А) от них произошли более высокоорганизованные папоротники
Б) они не дали начала более высокоорганизованным растениям
В) от них произошли более высокоорганизованные хвощи
Г) они произошли от одноклеточных водорослей
Ответ
620-4. Какой признак характерен для мхов?
А) придаточные корни развиваются от стебля
Б) споры образуются в коробочке
В) у них отсутствует побег
Г) опыление предшествует оплодотворению
Ответ
620-5. У мхов из споры развивается
А) коробочка на ножке
Б) семя
В) зелёная нить
Г) заросток
Ответ
620-6. Приспособленность мха сфагнума к жизни в условиях избыточного увлажнения проявляется в наличии
А) корневища с придаточными корнями
Б) клеток с хлоропластами
В) мертвых клеток
Г) ризоидов
Ответ
620-7. Представители какого отдела царства Растения изображены на рисунке?
Ответ
620-8. Какие растения относятся к отделу Мохообразные?
А) обитающие на суше и размножающиеся семенами
Б) листостебельные, не имеющие корней, размножающиеся спорами
В) все растения влажных мест обитания
Г) все травянистые растения
Ответ
620-9) Какие приспособления к поглощению большого количества воды появились в процессе эволюции у мхов?
А) ризоиды – выросты на стебле
Б) крупные мёртвые клетки
В) коробочки со спорами
Г) клетки тонкой покровной ткани
Ответ
620-10. У зелёных мхов, в отличие от водорослей,
А) клетки имеют большое и малое ядра
Б) оплодотворение происходит при наличии воды
В) слоевище разделено на ткани и органы
Г) осуществляется половое и бесполое размножение
Ответ
620-11. К какому отделу высших растений относится изображенное на рисунке растение?

А) Покрытосеменные
Б) Голосеменные
В) Папоротникообразные
Г) Мохообразные
Ответ
620-12. По каким признакам моховидных отличают от других растений?
А) в процессе их развития происходит чередование поколений
Б) размножаются спорами
В) имеют листья, стебель и ризоиды
Г) способны к фотосинтезу
Ответ
620-13. Папоротники, в отличие от зелёных мхов, имеют
А) ризоиды
Б) корни
В) листья
Г) стебли
Ответ
620-14. Из спор зелёного мха кукушкина льна развивается(-ются)
А) заросток в виде зелёной пластины
Б) предросток в виде зелёных нитей
В) растения с листьями
Г) семена будущего растения
Ответ
620-15. Из высших растений корней нет у
А) Цветковых
Б) Хвойных
В) Мхов
Г) Папоротников
Ответ
620-16. Папоротники распространены на Земле значительно шире мхов, так как они
А) имеют развитую корневую систему и более эффективно размножаются
Б) появились в ходе эволюции раньше и успели лучше приспособиться
В) широко выращиваются человеком для своих нужд
Г) успешно распространяются различными животными
Ответ
620-17. Наиболее простое строение среди высших растений имеют мхи, так как
А) у них нет корней
Б) стебель у них неветвящийся, с узкими листьями
В) они образуют органические вещества из неорганических
Г) у них есть воздухоносные клетки
Ответ
620-18. Почему мхи представляют тупиковую ветвь в эволюции растений?
А) они не освоили наземно-воздушную среду обитания
Б) они произошли от водорослей
В) они не имеют корней и размножаются спорами
Г) они не дали начало более высокоорганизованным растениям
Ответ
620-19. Представитель какого отдела царства Растения изображен на рисунке?

А) Папоротниковидные
Б) Голосеменные
В) Плауновидные
Г) Моховидные
Ответ
620-20. К какой группе организмов относят зеленые растения, не имеющие корней, размножающиеся спорами, в жизненном цикле которых преобладает половое поколение?
А) мохообразные
Б) папоротникообразные
В) голосеменные
Г) плауновидные
Моховидные — вечнозеленые, многолетние растения. Современная флора моховидных насчитывает около 25 000 видов. Тело моховидных представляет собой либо слоевище, прижатое к субстрату, либо стебель с листьями. У мхов корней нет, есть только ризоиды, выполняющие роль корней. У мхов слабо выражены проводящие, запасающие и механические ткани.
Фотосинтезирующее тело моховидных представляет собой гаметофит. На гаметофите развиваются мужские (антеридии) и женские (архегонии) половые органы. В антеридиях образуются двужгутиковые сперматозоиды, а в архегониях — крупные яйцеклетки. При помощи воды (в сырую погоду) сперматозоиды выходят из антеридиев, и один из них проникает в архегоний и оплодотворяет яйцеклетку. Из зиготы вырастает спорофит — бесполое поколение. Спорофит представлен коробочкой, сидящей на ножке. В коробочке образуются споры.
При прорастании споры формируется протонема — тонкая разветвленная нить. На протонеме образуются почки, которые дают начало новым гаметофитам. Гаметофиты способны к вегетативному размножению.
Во влажных, затененных местах и на болотах произрастает кукушкин лен, зеленые стебли которого образуют подушечки. Кукушкин лен — растение двудомное: на верхушке одних стеблей развиваются архегонии, а на других — антеридии. Оплодотворение происходит с помощью воды. После оплодотворения на женском растении развивается спорофит — коробочка на ножке. Спорофит желтоватого цвета, не способен к фотосинтезу и питается за счет гаметофита. В коробочке созревают споры, из которых после прорастания образуется нитчатая протонема. На протонеме развиваются почки, а из них — новые зеленые гаметофиты.
Сфагновые мхи широко распространены в умеренных и холодных широтах. Они образуют сплошной покров на болотах и во влажных лесах.
Стебли у сфагнума прямостоячие, листья расположены мутовчато, стебель ветвится, ветви несут листья. Листья однослойные, состоят из клеток двух типов — живых зеленых, которые содержат хлоропласты, и бесцветных мертвых. Оболочки мертвых клеток имеют крупные отверстия, через которые в них проникает вода, поэтому сфагнум накапливает воду.
Сфагнум — растение однодомное: на одном стебле образуются архегонии и антеридии. Оплодотворение происходит с помощью воды, по которой сперматозоид подплывает к яйцеклетке. Из зиготы формируется спорофит. Он представляет собой коробочку на ножке. Спорофит — нефотосинтезирующее поколение, он питается за счет зеленого гаметофита. В коробочке созревают споры. Они высыпаются и распространяются с помощью ветра или воды. Из споры прорастает протонема, на которой развиваются стебли гаметофита.
Сфагнум не имеет ризоидов, он впитывает воду всей поверхностью тела. Нижняя часть стебля сфагнума погружена в воду и постепенно отмирает. Отмершие части стебля опускаются на дно водоема, где медленно разлагаются практически без доступа кислорода. В результате образуется торф. Процесс торфообразования протекает очень медленно — 1 см за 10 лет. Сфагнум содержит карболовую кислоту, обладающую бактерицидными свойствами, поэтому в торфяных отложениях хорошо сохраняются останки животных и растений.
Торф — ценное удобрение, также используется в химической промышленности как сырье для получения лаков, красок, пластмасс, метилового спирта, карболовой кислоты и в качестве топлива.
Значение мхов в природе в основном связано со сфагновыми мхами, вызывающими быстрое заболачивание и торфообразование.
Издали мхи напоминают ковёр или мех зеленоватого, красного или бурого цвета. Вблизи можно увидеть, что ворсинки ковра — это стебельки с листочками. Листочки мхов состоят обычно из одного слоя клеток. Стебли мхов выносят листья к свету. У многих мхов на стеблях есть тонкие выросты — ризоиды, которыми они цепляются за грунт.
Зелёный мох кукушкин лён — небольшое растение, обычно не более 20 см. У кукушкина льна стебли буровато-зелёные, неветвящиеся, густо покрыты узкими листьями.
Мхи растут густыми дернинками. Дождевая вода помогает попасть сперматозоидам на верхушки женских растений. Проникают к яйцеклеткам, происходит оплодотворение, и образуется зигота. На следующий год из зиготы развивается коробочка со спорами. Спора попадает на влажную почву и прорастает, образуя тонкую зелёную нить. Нить ветвится; на ней появляются почки, из которых вырастают побеги мха.
Что делаем. Рассмотрите растение зелёный мох кукушкин лён.
Найдите: стебель и листья, ризоиды, (тонкие бурые выросты), ножку и коробочку (на верхушке стебля).
Рассмотреть лист под микроскопом и зарисовать в тетради.

Определить форму стебля (ветвистая, неветвистая).

Определить мужское это, или женское растение.

Под лупой рассмотреть продольный разрез коробочки с крышечкой и внутри найти споры. Рассмотреть споры под лупой.
Общая характеристика мхов. Моховидные - обширная группа высших растений, очень различающихся по внешнему строению. Во всем мире их насчитывается около 25 тыс. видов. Среди высших растений по количеству видов они занимают второе место после цветковых.
Моховидные представляют собой очень древнюю группу в царстве растений. Почти все они - многолетние растения. Обычно мхи низкорослы: их высота колеблется от нескольких миллиметров до 20 см. Они всегда растут в местах повышенной влажности.
Средимоховидных выделяют два больших класса - Печеночники и Листостебельные мхи .
У печеночников тело представлено разветвленным зеленым плоским слоевищем. У листостебельных мхов хорошо видны стебли и мелкие зеленые листья, т. е. имеются побеги. Те и другие имеют ризоиды , которые поглощают воду из почвы и закрепляют растения. Все моховидные характеризуются значительной простотой внутреннего строения. В их теле имеются основная и фотосинтезирующая ткани, но проводящие, механические, запасающие и покровные ткани отсутствуют.
Печеночники - очень древние мхи. Они особенно богато представлены в тропиках. Один из распространенных видов печеночников - маршанция (рис. 95), обитающая во влажных местах, не занятых травой. У нее стелющееся листовидное слоевище, ризоидами прикрепленное к почве. В слоевище наблюдается разделение ткани на основную (в нижней части тела) и фотосинтезирующую (в верхней части тела). К маршанциевым относится теплолюбиваяриччия водная, разводимая аквариумистами.
Маршанция
Листостебельные мхи в растительном покрове Земли играют значительно большую роль, чем печеночники. Один из наиболее известных зеленых листостебельных мхов - кукушкин лен , илиполитрихум обыкновенный (рис. 96), часто встречается в хвойных лесах, около сфагновых болот, в сырых местах. Многолетние крупные растения этого вида (9-17 см. в длину), растущие группами, нередко покрывают обширные площади в лесной зоне и в тундре.
Кукушкин лен
Размножение моховидных. Моховидные размножаются бесполым и половым путем. Бесполое размножение осуществляется спорами (поэтому их относят к споровым растениям) и вегетативным путем (частями слоевища, стеблей, листьев), а половое - с помощью гамет.
Для полового размножения у мхов развиваются специальные органы, в которых образуются гаметы: мужские - сперматозоиды и женские - яйцеклетки. У слоевищных мхов половые органы располагаются на верхней поверхности слоевища, а у листостебельных - в верхушечной части побегов.
В водной среде с помощью жгутиков сперматозоиды передвигаются к яйцеклетке и оплодотворяют ее. Без воды сперматозоиды не могут достигнуть яйцеклетки. После оплодотворения из появившейся зиготы развивается особый орган - коробочка , в которой формируются споры. С помощью спор моховидные размножаются и расселяются. Из споры вначале развивается многоклеточная тонкая зеленая нить -протонема . На ней вскоре из почек формируются пластинчатые талломы или облиственные побеги.
У кукушкина льна образующаяся из зиготы коробочка расположена на длинной жесткой ножке, имеет крышечку и покрыта колпачком. При созревании спор крышечка коробочки открывается и споры высыпаются. Они очень мелкие и легкие, поэтому далеко разносятся. Чем длиннее ножка, тем дальше могут рассыпаться споры. Попав в благоприятные условия, споры прорастают, формируют протонемы, и весь цикл развития этого мха повторяется вновь.
Значение мхов.
Появление на почве кукушкина льна - сигнал, предупреждающий о возможном заболачивании почвы. Кукушкин лен может создавать большие и плотные покровы почвы, что способствует накоплению воды. Как мощный накопитель влаги он способствует возникновению болот. В местах поселения кукушкина льна, накапливающего влагу, вскоре может поселиться сфагнум.
В отличие от кукушкина льна и других зеленых мхов сфагнум иногда называют в народе белым мхом (рис. 97). В листьях сфагнума хлорофиллоносные клетки чередуются с большими мешковидными клетками, заполненными воздухом или водой.
Сфагнум способен быстро накапливать в теле много капельно-жидкой воды и тем способствовать заболачиванию почвы.

Число видов 13 500 (80 семейств и 657 родов). Распространены более широко, чем сфагновые мхи, встречаясь в разнообразных экологических условиях от тундры и лесотундры до степей и пустынь. Наиболее типичные местообитания зеленых мхов, где они подчас явно доминируют или формируют сплошной покров,- тундры, болота и некоторые типы лесов. Каждому специфическому местообитанию, например: определенному типу леса, свойственны свои виды. Для дубрав и смешанных лесов Восточной Европы очень характерны виды родов мниум - Mnium и климациум - Climacium, в более северных, менее увлажненных лесах преобладают виды родов плеуроциум - Pleurozium, дикранум - Dicranum, хилокомиум - Hylocomium. В хвойных лесах более высоких широт на первый план выступают виды рода кукушкин лен - Polytrichum. Зеленые мхи по сравнению со сфагновыми отличаются большим разнообразием строения.
Рассмотрим строение одного из наиболее широко распространенных зеленых мхов - кукушкина льна (Polytrichum commune). Стебелек, плотно покрытый филлоидами, чаще всего не ветвится и достигает 15 см высоты, а иногда и более. По сравнению со сфагновыми мхами гаметофит в структурном и функциональном отношениях более совершенен и разнообразен (рис. 199). Осевая часть (стебелек) гаметофита имеет в центре более вытянутые клетки, которые соответствуют ксилеме и флоэме, что облегчает проводящую функцию. Настоящего проводящего пучка нет. Филлоид состоит из нескольких слоев клеток и более дифференцирован. В его срединной части обособляются удлиненные проводяшие и толстостенные механические клетки, аналогичные средней жилке настоящего листа. На верхней поверхности филлоида образуется зеленая бахрома из коротких хлорофиллоносных нитей (ассимиляторов) - фотосинтезирующая ткань. У основания стебелька развиваются многоклеточные ризоиды - аналоги корней. Органы полового размножения расположены на разных особях - мужских и женских гаметофитах. Строение антеридиев и архегониев не имеет принципиальных отличий с: таковых у сфагновых мхов.
Спорогон состоит из гаустории, ножки и коробочки. Однако по сравнению со сфагновыми мхами части спорогона более развиты, особенно коробочка. Основные отличия строения коробочки следующие: а) верхняя часть ножки сильно разросшаяся и содержит хлорофиллоносную ткань; б) наверху коробочки до созревания спор находится волосистый колпачок, развивающийся из стенки брюшка архегоння; в) внутри расширенной части коробочки - урночки- расположена колонка, где она расширяется и формирует эпифрагму - тонкостенную перегородку, над которой находится крышечка, г) спорангий представляет собой цилиндрический мешок, расположенный вокруг колонки; он прикреплен к стенке урночки и к колонке особыми нитевидными образованиями; д) коробочка имеет специальное приспособление для рассеивания спор - перистом. Он представляет собой ряд зубчиков, расположенных по краю урночки.
Между зубчиками перистома есть сквозные поры, через которые в сухую погоду высыпаются споры. Из споры вырастает протонема в виде зеленой ветвящейся нити. На протонеме возникают особые почки, из которых со временем развивается взрослый гаметофит.
199. Кукушкин лен - Polytrichum commune. A - мужской гаметофит; Б - верхушка мужского
гаметофита (продольный срез); В - женский гаметофит; Г - верхушка женского гаметофита (продольный срез); Д - стебель (поперечный срез); Е - филлоид общий вид и поперечный срез); Ж - спорогон, развившийся на женском гаметофите;
3 - коробочка спорогона (с колпачком, без колпачка и продольный срез)
1:антеридий, 2 - парафиза, 3 - архегоний, 4 - эпидерма, 5 - «кора», 6 - клетки, выполняющие - функцию флоэмы, 7 - клетки, выполняющие функцию ксилемы, 8 - паренхимные клетки, 9 - механические клетки, 10 - ассимиляторы, // - ризоиды, 12 - колпачок, 13 - крышечка, 14 -- эпифрагма, 15 - стенка урночки, 16 - колонка, 17 - спорангий, 18 - апофиза, 19 - ножка.
Происхождение мохообразных еще не выяснено.
1.Одни авторы филогенетических систем выводят их из риниеобразных. Они основываются на сходстве структуры спорангия и устьиц риниеобразных и сфагновых мхов. Более примитивное строение мохообразных объясняют как:вторичное в связи с возвратом к водным местообитаниям.
2.Более мотивирована концепция другой группы ученых: мохообразные происходят непосредственно от водорослей. В этом случае предполагается, что риниеобразные и папоротникообразные также возникли от водорослей, и представляют собой особую параллельную ветвь развития, где прогрессировал другой тип чередования фаз в жизненном цикле с доминированием спорофита. Предковыми формами этой линии, которая оказалась исключительно прогрессивной, по-видимому, были водоросли, у которых спорофаза и гаметофаза представлены разными индивидуумами, развивающимися независимо друг от друга и не различающимися между собой и морфологически.
ОТДЕЛ ПЛАУНООБРАЗНЫЕ - LEPIDOPHYTA
Плаунообразные представляют мелколистную линию эволюции. Появление их датируют силурским периодом палеозойской эры. Первые плаунообразные были травянистыми растениями, затем возникали одревесневающие формы. Ныне живущие плаунообразные вместе со многими известными ископаемыми видами представляют значительное разнообразие жизненных форм. Среди них встречаются деревья, кустарники, травы. Органы спорофита - корень, стебель и мелкие, иногда чешуевидные листья, слабо дифференцированные, с одной - двумя неветвящимися жилками. Они представляют собой по существу выросты стебля - филлоиды. Поэтому побеги плаунообразных не имеют хорошо выраженных узлов и междоузлий. Стебли и корни характеризуются ди-томическим ветвлением. Спорангии или одиночные, расположенные на верхней стороне листьев (спорофиллюв), или собраны в колоски. Древовидные плаунообразные были широко и обильно распространены в середине каменноугольного периода, а затем вымерли. Травянистые формы оказались более пластичными и дошли до наших дней. Отдел Плаунообразные чаще всего подразделяют на 3 класса: Плауновидные, Чешуедревовидные и Полушниковидные.
КЛАСС ПЛАУНОВИДНЫЕ - LYCOPODIINAE
Род плаун - Licopodium включает 400 видов. На территории России представлен 14 видами. Наиболее широко распространен плаун булавовидный - L. clavatum. Встречается преимущественно в хвойных лесах. Из других видов можно отметить плаун-баранец - L. selago (лиственные леса, альпийские и субальпийские луга), плаун обоюдоострый - L. anceps (сосновые леса).
Рассмотрим плаун булавовидный - L. clavatum
Рис. 127. Жизненный цикл плауна булавовидного - L усо podium clavatum: £--деление зиготы, 2 - зародыш спорофита, 3- взрослое растение спорофита, 4 - спорофилл со спорангием, 5 - спорогенная клетка, 6-8 - развитие спор из спорогенной клетки путем мейоза, 9 - спора, 10 - прорастание споры, образование протонемы, // - гаметофит с архегониями и анте-ридиями, 12 - архегоний с яйцеклеткой, 13 - антеридий со сперматозоидами, 14 - сперматозоид,
15 - оплодотворение
Спорофит представлен многолетним вечнозеленым растением. Стебель стелется по земле и дает вертикальные, дихотомически ветвящиеся побеги до 25 см высоты. Побеги заканчиваются спороносными колосками или верхушечными почками. От лежачего стебля отходят дихотомически ветвящиеся корни. Стебель и ветви густо покрыты спирально расположенными мелкими ланцетно-линейными листьями. На поперечном сечении стебля можно рассмотреть структуру стелы (центрального цилиндра), напоминающую концентрический проводящий пучок, и кору. Вторичный рост отсутствует. Коровую часть пронизывают ответвления стелы (листовые следы), которые переходят в листья и формируют их среднюю жилку. Стебель и листья покрыты эпидермой с хорошо развитыми устьицами. Спороносные колоски на длинных ножках, на их оси сидят спорофиллы, со спорангиями на верхней стороне. После редукционного деления материнских клеток образуются гаплоидные споры. Все споры по форме и величине одинаковы. Споры содержат до 50% жирного невысыхающего масла. Споры попадают на землю и на глубине нескольких сантиметров из них медленно, в течение 12- 20 лет, развивается гаметофит. По форме, он напоминает луковичку, позднее разрастается до 2-3 см в диаметре и имеет блюдцевидную форму. Гаметофит имеет ризоиды, но лишен хлоропластов. Клетки, расположенные под эпидермой, содержат мицелий гриба (симбиоз). Однако у некоторых видов гаметофит развивается на поверхности почвы, и тогда в его клетках появляются хлоропласты. Антеридии и архегонии расположены рядом на верхней стороне и погружены в паренхимную ткань. Антеридии овальной формы, архегонии - колбовидной. Сперматозоиды многочисленные, двужгутиковые. Из зиготы развивается зародыш. Зародыш спорофита вначале внедрен в ткань гаметофита и в какой-то мере питается за его счет. Вскоре его корни проникают в почву, и начинается долгая самостоятельная жизнь спорофита.
Хозяйственное значение плаунов невелико. Они не поедаются животными. Издавна используются лишь споры плаунов, содержащие невысыхающее масло. Их применяют в медицине в качестве детской присыпки. Иногда споры плауна используют при фасонном литье для обсыпания стенок моделей, чтобы отливаемая деталь легко отделялась от формы.
КЛАСС ХВОЩЕВИДНЫЕ - EQUISETINAE
В современной флоре класс представлен одним семейством Хвощевые - Equisetaceae. В составе семейства только один род хвощи. Другие роды семейства давно вымерли. Общее число видов 30- 35. Распространены они на всех континентах мира (кроме Австралии) до- .вольно широко, преимущественно в условиях влажных местообитаний. В России 13 видов. Современные хвощи - сугубо травянистые растения, хотя некоторые из них достигают значительных размеров (12 м). Это лазающие растения тропического леса с тонкими стеблями. Большинство хвощей имеет однолетние надземные побеги, только немногие вечнозеленые (в лесной зоне широко распространен вечнозеленый хвощ зимующий - Е. hiemale). Стенки клеток эпидермы часто инкрустированы кремнеземом, что обесценивает кормовое значение хвощей.
Подземная часть представлена сильно развитым корневищем. Боковые ветви корневища служат местом отложения запасных веществ и у некоторых превращаются в клубни, которые служат также и органами вегетативного размножения. От горизонтального корневища отходят надземные побеги. Все побеги и корневища нарастают верхушкой.У хвоща полевого рано весной развиваются неветвящиеся бесхлорофилльные побеги. На верхушке они несут спороносные колоски (рис. 202, Б).Спорангии расположены на особых спорангиофорах 1 (рис. 202, В) У современных хвощей все споры морфологически одинаковы и обоеполы. После созревания спор спороносные побеги отмирают. Зеленые вегетативные побеги типичным мутовчатым ветвлением появляются позднее и вегетируют до поздней осени, но развиваются из того же корневища, что и спороносные (рис. 202, А). Снабженные элатерами споры хвоща полевого прорастают на почве в хлорофиллоносные гаметофиты в виде лопастных пластинок, морфологически различных. Одни из них - мужские с антериями, формирующими многожгутиковые сперматозоиды, другие -женские, с архегониями. Последние более рассеченной формы(202, Д) Некоторые виды хвощей имеют важное значение как кормовые растения: хвощ ветвистый - Е. ramosissimum, хвощ пятнистый - Е. variega-~:, хвощ зимующий - Е. hiemale. Встречаются и ядовитые виды: 
Рис. 202. Хвощ полевой - Equisetum arvense. A - вегетативный побег; Б - спороносный побег; В - спорангиофор со спорангиями (внешний вид и поперечный срез); Г -спора;
Д - гаметофиты (мужской и женский):
/ - корневище, 2 - спороносный колосок, 3 - щиток, 4 - спорангий, 5 - ножка, 6 - перина, 7 - элатеры, 8 -> антеридий, 9 - архегоний
хвощ болотный - Е. palustre, хвощ дубравный - Е. nemorosum и др. Хвощ полевой - Е. arvense нередко встречается в посевах как злостный сорняк, причем его кормовые качества оспариваются.
ОТДЕЛ ПАПОРОТНИКООБРАЗНЫЕ - PTEROPHYTA
Общее число видов около 15 тыс. Это наиболее обширный отдел среди высших споровых. Распространены на всех континентах мира, но преимущественно в тропиках. Многие виды вымерли. Первые папоротникообразные представлены главным образом деревьями, иногда крупными. В тропических лесах Австралии и Новой Зеландии в наши дни произрастают древовидные папоротникообразные, достигающие 20 м высоты. Стволы древовидных видов колонновидные, не ветвящиеся, как у некоторых покрытосеменных, например у пальм. Общее направление эволюции папоротникообразных - от древовидного прямостоячего радиально-симметричного тела спорофита к травянистому простертому - дорзовентральному
Таблица 3.7. Систематика и основные признаки Pteridophyta
Отдел Pteridophyta (папоротникообразные)
Чередование поколений, при котором доминирует спорофитное поколение Гаметофит редуцирован до крошечного, простого заростка У спорофита есть настоящие корни, стебли и листья с проводящими тканями
Класс Lycopsida (плауновидные)-плауны
Класс Sphenopsida (клинолистовидные, членистые) - хвощи
Класс Pteropsida (папоротниковидные) -папоротники
Листья сравнительно небольшие (микро-филлы) 1 * и расположены спирально вокруг стебля
Равноспоровые и разноспоровые формы Спорангии обычно в спороносных шишках (стробилах)
Примеры: Selaginella - разноспоровый плаун
Lycopodium - равноспоровый плаун
Листья сравнительно небольшие (микрофиллы) 1 * и расположены мутовками вокруг стебля
Равноспоровые
Спорангии в спороносных шишках
(стробилах) на хорошо заметных
спорангиофорах
Примеры: Equisetum (единственный род, сохранившийся до наших дней)
Листья сравнительно крупные (макрофиллы) 1 \ называются вайями и расположены спирально вокруг стебля
Равноспоровые (преимущественно) Спорангии обычно собраны в кучки (сорусы)
Пример: Dryopteris filixmas (мужской папоротник, щитовник) Pteridium (папоротник-орляк)
Микрофиллы - листья, имеющие одиночную центральную жилку. Обычно небольшие. Макрофиллы-листья имеющие разветвленные жилки. Крупные.

Рис. 203. Выводковые почки Папоротникообразных - Pterophyta. Л - цистоптериса луковичного - Cystopteris bulbifera; Б - асплениума - Asplenium viviparum: I - выводковые почки, 2-3 - фазы прорастания выводковых почек
Редукция вегетативного тела произошла вследствие приспособления к эпифитному и геофитному образу жизни в условиях умеренного и холодного континентального климата.
От предыдущих высших споровых папоротникообразные отличаются крупнолистностью (мегафиллией). Однако у них листья особого рода. Они длительно нарастают верхушкой, а не основанием, выводковые почки закладываются в мезофилле листа (рис. 203). Есть все основания считать листья папоротникообразных гомологами стеблей (кладодиями). Поэтому их часто называют вайями, или плосковетками. На нижней стороне вай расположены спорангии. Чаще всего они группируются кучками - сериями. Тогда спорангии бывают покрыты покрывальцем -индузием (см. рис. 207). Иногда же спорангии свободно рассеяны на нижней поверхности листьев. Полагают, что изначально вайя была одновременно и фотосинтезирующим, и спороносным органом. Позднее произошла дифференциация: верхние вайи специализировались для спороношения (см. рис. 208). В других случаях наблюдается специализация даже отдельных участков одной вайи(осмунда). Стробил - спороносный колосок, столь характерный для предыдущих споровых, у лептоспорангиатных папоротников отсутствует. Спорофит -сложно устроенное растение. От корневища почти вертикально отходят вайи, а вниз- придаточные корни, которые, как и корневище, состоят из хорошо дифференцированных тканей.
Споры, полученные в результате редукционного деления (п хромосом), рассеиваются благодаря разрыву стенки спорангия. Число спор на одном растении исчисляется десятками и сотнями миллионов, а иногда и миллиардами. Гаметофит обоеполый, изредка раздельнополый, например у водных папоротников. Форма гаметофита довольно однообразна: у наших видов чаще всего сердцевидная (см. рис. 207), у тропических - в виде ветвистой пластинки или нитевидная. Из зиготы формируется зародыш спорофита (2 п хромосом). Зародыш (рис. 204), кроме обычных частей, имеет гаусторию - ножку, которой он внедряется в ткани гаметофита и поглощает пищу.

Рис. 37. Зародыш спорофита адиантума - род Adianthum (поперечный срез):
/ - пластинка гаметофита, 2 - ризоиды, 3 - неоплодотворенные архегонии, 4 -
гаустория зародыша, 5 - зародышевый корешок, 6 - первый лист зародыша
Зародышевый корешок вскоре редуцируется, и на смену ему возникают придаточные корни.
Гаметофит приспособлен к жизни в условиях обильной влаги (как водоросли), спорофит - сухопутное растение. Некоторое исключение составляет спорофит водных папоротников. Таким образом, приспособительная эволюция папоротникообразных к сухопутным условиям шла по линии спорофита. Однако папоротникообразные не могли завоевать сушу, поскольку гаметофит остановился на уровне водорослевого периода существования. Половой процесс у них, как у водорослей и мохообразных, неминуемо связан с водной средой.
Папоротникообразные подразделяют на 3 класса: Первопапоротники, Настоящие папоротники, Лептоспорангиатные папоротники.
КЛАСС ПЕРВОПАПОРОТНИКИ - PRIMOFILICIDAE
Разнообразная, но слабо изученная группа ископаемых папоротникообразных. Общее число видов 60. Основные черты строения: дихотомически ветвящиеся стебли; спорангии возникают из группы клеток на концах веточек, как у риниеобразных, и лишены индузия; стенка спорангия многослойная, без кольца утолщения, способствующего разрыву и рассеиванию спор. Наиболее примитивные похожи на риниеобразных, которые вероятнее всего и являются родоначальниками папоротникообразных.
КЛАСС НАСТОЯЩИЕ ПАПОРОТНИКИ - EUFILIС IDAE
Из числа современных папоротникообразных это наиболее древняя и наиболее примитивная группа. Здесь объединены виды, которые имеют спорангии с многослойной стенкой, как первопапоротники. Наибольшего расцвета они достигли в палеозойскую эру. Сейчас это уже угасающая ветвь развития. Общее число видов около 300. Класс включает 2 порядка: Ужовниковые и Мараттиевые.
Порядок Ужовниковые - Ophioglossales с одним семейством Ужовниковые - Ophioglossaceae, которое содержит 3 рода. Общее число видов около 90. Жизненные формы: травянистые растения (иногда вечнозеленые), эпифиты. Распространены по земному шару в тенистых лесах, но приурочены к северной умеренной зоне (род ботрихиум) или тропической (роды ужовник и гельминтостахис).
Типичный представитель класса - ужовник обыкновенный - Ophioglossum vulgatum(рис.205,А). Встречается, хотя и изредка, но почти повсеместно в лесах, на сырых местообитаниях от Северной Африки до Скандинавии. 
Рис. 205. Ужовник обыкновенный - Ophioglossum vulgatum (Л); гельминтостахис цейлонский- Helminthostachys zeylonica (Б): 1 - спорангий
В России преимущественно по мшистым луговинам. Это небольшое растение до 10-15 см высоты с почти вертикальным корневищем. Важно отметить один из признаков сходства с уже рассмотренными высшими споровыми растениями: наличие простого или ветвистого «спороносного колоска». Колосок этот на более или менее длинной ножке. У ужовниковых развиты придаточные корни, обычно мясистые, лишенные корневых волосков (микориза). По-видимому, ужовниковые - прямые потомки первопапоротников.
Порядок Мараттиевые - Marattiales с одним семейством Мараттиевые - Marattiaceae и с шестью родами. Много ископаемых видов известно из каменноугольного периода палеозойской эры. Распространение - влажные тропические леса обоих полушарий. Это многолетние растения, то мелкие травянистые, то крупные одревесневшие. Стебли прямостоячие, неветвящиеся, иногда сильно укороченные. Иногда же стебель сильно редуцирован и имеет вид дорзовентрального, слабо ветвящегося корневища. Вайи перистые, реже цельные. В отдельных случаях они достигают огромной величины - 4-5 м (роды макроглоссум, мараттия и др.)« Спорангии в сориях, у более современных форм они срастаются в синан-гии \ Стенка спорангия во всех случаях многослойная. Гаметофиты наземные, многолетние, крупные - до 3 см.
КЛАСС ЛЕПТОСПОРАНГИАТНЫЕ ПАПОРОТНИКИ - LEPTOFILICIDAE
Класс объединяет подавляющее большинство современных папоротникообразных. Лептоспорангиатные папоротники представляют значительное разнообразие жизненных форм: древовидные, лианы, травянистые эпифиты (влажные тропические леса), многолетние корневищные травы (умеренная и холодная зоны). Особенно велико значение их в сложении растительных сообществ тропического леса. Однако и в умеренной зоне, особенно во влажных местообитаниях, лептоспорангиатные папоротники иногда играют видную роль. Подавляющее большинство видов равноспоровые, вполне сухопутные растения (спорофит!). Они формируют порядок Собственно папоротники (около 10 тыс. видов). Остальные (около 120 видов) - разноспоровые, водные и водно-болотные растения объединены в порядок Водные папоротники.
Порядок Собственно папоротники - Filicales. Большинство - многолетние корневищные травы. В тропической флоре встречаются прямостоячие деревья и лианы. Общие признаки с
ледующие: а) ярко выраженная

Рис. 206. Орляк - Pteridium aquilinum. A - спорофит; Б - часть вайи с сорпями; В - часть корневища; Г - поперечный срез корневища (схема); Д - проводящий иучт
(поперечный срез):
/ - кора (наружная и внутренняя), 2 - механическая ткань, S -проводящий пудок, 4 - эндодеркж, 5 - перицикл, 6 - флоэма» 7 - ксилема

э ис. 207. Мужской папоротник - Dryopteris fillx-mas. A - спорофит; Б - часть вайи: сориями; В - поперечный срез вайи с сорием; Г - спорангии; Д - спора; Е - протонема; Ж - гаметофит; 3-антеридий; И - архегонии; /С-молодой спорофит:
1 - плацентал 2 * * ножка спорангия, 3 - спорангий, 4 - индузий, 5 -* кольцо утолщения, 6 - ризоиды.
7 - антеридии, 8 = архегонии
крупнолистность - мегафиллия, причем вайи, несомненно, стеблевого происхождения; б) у большинства внетропических видов стебли распростертые, укороченные, с хорошо выраженным дор-зовентральным строением: вайи развиваются исключительно на верхней стороне, а придаточные корни - на нижней; в) обычно стебли погружены в почву и более или менее преобразованы в корневища; г) зрелые спорангии имеют оболочку из одного слоя клеток и кольцо утолщения, способствующее ее разрыву; д) спорангии образуются на нижней стороне ассимилирующих вай, иногда на особых спорофиллах, но никогда не бывают собраны в спороносные колоски; е) гаметофиты обычно состоят из одного слоя клеток, наземные, хлорофил-лоносные.
На территории СССР известно около 120 видов этого порядка. Многие из них широко распространены. Некоторые же виды, например орляк - Pteridium aquiliпит, нередко выступают в качестве доминирующих растений в светлых лесах (рис. 206). Вайи на
длинных черешках, крупные, одиночные, наверху тройчаторассеченные. Корневища мощные, содержатбольшое количество запасных веществ, простираются горизонтально на значительной глубине (до 25-30 см). Осевые органы орляка, как и корневище, имеют сложное микроскопическое строение в связи с хорошо дифференцированными тканями и широким разнообразием гистологических элементов. Широко распространен во влажных лесах папоротник мужской-Dryopteris filix-mas (рис. 207). Оригинальным строением из-за специализации вай отличается страусник - Matteuccia struthiopteris (рис. 208).
Это один из наиболее декоративных папоротников в нашей флоре.
Порядок Водные папоротники - Hydropteridales. В порядке объединено 3 семейства: Марсилиевые - Marsiliaceae, Сальвиниевые - Salvi-niaceae и Азолловые - Azollaceae. Общее число видов ПО. Это растения преимущественно водно-болотных местообитаний тропических и субтропических лесов. В дельте Волги, на Северном Кавказе, в Карпатах встречается представитель первого семейства - марсилия четырехлистная - Marsilia quadrifolia. Еще более продвинута на север и часто встречается сальвиния плавающая - Salvinia natans из семейства Сальвиниевые. Характерная их особенность - разноспоровость и соответственно наличие микро- и мегаспорангиев. В мегаспорангии находится по одной мегаспоре, в микроспорангиях образуется много микроспор. Микро- и мегаспорангии развиваются в спорокарпиях, аналогичных сориям. У водных папоротников наблюдается дальнейшая редукция гаметофитов. Так, вегетативная часть мужского гаметофита представлена лишь двумя клетками. Женский гаметофит настолько мал, что значительная часть
Рис. 208. Страусник обыкновенный - Matteuccia struthiopteris. A - спорофит; Б -
часть вайи с сорием; В - срез сория: 2 - стерильный лист, 2 - спорфилл, 3 - спорангии
его помещается в оболочке мегаспоры. На первых фазах, пока еще не сформировались вегетативные органы, спорофит питается за счет зеленого женского гаметофита. Тесная взаимосвязь мегаспоры, женского гаметофита и нового молодого спорофита водных папоротников проливает свет на происхождение семян.
Значение папоротникообразных в природе велико. Нередко они выступают в роли важнейшего компонента многих растительных сообществ, особенно тропических и субтропических лесов, а также северных, преимущественно широколиственных. Папоротникообразные - один из важных объектов декоративного цветоводства. В условиях закрытого грунта, оранжерей и комнатного озеленения культивируют особенно эффектный эпифит - платицериум (род Platicerium) и целый ряд корневищных видов: птерис (род Pteris), асплениум (род Asplenium) и др. В открытом грунте наиболее обычны чистоуст (род Osmunda) и страусник (род Matteuccia).
Голосеменные - древние растения, их ископаемые остатки находят в слоях девонского периода палеозойской эры. Многие виды и даже целые порядки, например семенные папоротники, кордаиты, известны исключительно как ископаемые. В настоящее время в мировой флоре насчитывается около 600 видов голосеменных. Это преимущественно деревья, реже одревесневающие лианы или кустарники. Травянистые формы отсутствуют. Ветвление в основном моноподиальное. Стебель имеет вторичное утолщение, сосудов нет, древесина состоит только из трахеид. Голосеменные делятся на две группы: одни имеют крупные рассеченные листья, похожие на листья пальм, или папоротников, другие - мелкие, цельные, чешуевидные или игольчатые (хвоя). Голосеменные за небольшим исключением - вечнозеленые растения. Корни - главный и боковые, с микоризой. Один из наиболее важных признаков всех голосеменных- наличие семязачатков (семяпочек). Семязачаток представляет собой мегаспорангий, окруженный особым защитным покровом - интегументом. Семязачатки расположены открыто, на мегаспорофиллах, и из них после оплодотворения развиваются семена. Сравнительный анализ соотношения в жизненном цикле гаплоидной (гаметофит) и диплоидной (спорофит) фаз, а также структуры органов полового размножения и хода полового процесса у голосеменных и смежных с ними групп растений дает представление о степени родства голосеменных и папоротникообразных, с одной стороны, и голосеменных и покрытосеменных - с другой.
Рассмотрим особенности размножения голосеменных на примере сосны обыкновенной - Pinus sylvestris. Спорофит - это дерево до 50 м высоты, достигающее возраста 400 лет. Побеги двух видов - удлиненные и укороченные. Спорообразование начинается примерно на 30-40-м году жизни. Спорофиллы собраны в шишки двух видов, резко различающиеся между собой, но образующиеся на одном растении: мужские, расположенные группами, и женские - одиночные (рис. 43).
Мужская шишка (длина 4-5 мм, ширина 3-4 мм) возникает в пазухе чешуйки, на месте укороченного побега. Она представляет собой побег с хорошо развитой осью (стержнем), на которой спирально расположены микроспорофиллы - редуцированные спороносные листья. У основания оси имеются чешуйки, играющие защитную роль. Микроспорофиллы яйцевидной формы, тонкие, плоские, с двумя микроспорангиями (пыльниками) на нижней стороне. К осени в микроспорангии заканчивается обособление

Рис. 43. Сосна обыкновенная - Pinus sylvestris. A - ветка спорофита с шишками; Б - молодая женская шишка (часть продольного среза); В - семенная чешуйка с семязачатками; Г - семязачаток (продольный срез); Д - мужская шишка (часть продольного среза); Е - микроспорофилл (продольный срез); Ж-пыльцевое зерно; 3 -
семя (продольный срез); И-семенная чешуйка зрелой шишки:
/ - группа мужских шишек, 2 - молодая женская шишка, 3 -» зрелые женские шишки; 4 - интегу-мент, 5 - микропиле, 6 - нуцеллус, 7 - эндосперм (женский гаметофит), 8- архегоний, 9 - пыльцевая трубка со спермиями, 10- микроспорангий, // - экзина, 12- интина, 13 - воздушный мешок, 14 - вегетативная клетка, 15 - антеридиальная клетка, 16 - проталлиальные клетки, 17 - кожура семени, 18 - зародышевый корешок, 19 - семядоли, 20 - гипокотиль
материнских клеток микроспор. Они окружены тапетумом. Весной происходит редукционное деление. В результате каждая диплоидная материнская клетка образует четыре микроспоры (рис. 44). Микроспора одноядерна, одета двумя оболочками - интиной и экзиной. Она несет два сетчатых воздушных мешка, которые возникают вследствие расхождения экзины и интины. Здесь же, в микроспорангии, происходит прорастание микроспоры и развитие мужского гаметофита, называемого пыльцой. Он развивается внутри оболочек микроспоры и редуцирован в еще большей мере, чем у рассмотренных ранее разноспоровых растений. Сначала в результате деления ядра микроспоры образуются две проталлиальные клетки, которые вскоре исчезают. Эти эфемерные клетки - единственные вегетативные клетки заростка. Затем ядро микроспоры делится еще раз, и вновь появляются две клетки: антеридиальная и вегетативная. Мужской гаметофит полностью лишен антеридиев. Покровы оболочки микроспоры становятся покровами пыльцы. Ко времени ее созревания микроспорангии вскрываются продольной щелью и пыльца высыпается наружу. Воздушные мешки облегчают перенос пыльцы ветром. Дальнейшее развитие мужского гаметофита происходит после опыления, т. е. на женских шишках внутри семязачатка. Развитие микроспор и структура пыльцы у всех голосеменных более или менее однообразны.
Женские шишки образуются на верхушках молодых побегов. Строение их более сложно и они относительно больших размеров, чем мужские шишки

Рис. 44. Развитие мужского гаметофита (пыльцы) сосны обыкновенной - Pinus sylvestris. A - деление материнской клетки; Б - тетрада микроспор; В - микроспора; Г - Е - образование мужского гаметофита (пыльцы); Ж-прорастание пыльцы:
/-2 - проталлиальные клетки, 3 - антеридиальная клетка, 4 - вегетативная клетка, 5 -клетжа-ножка, 6 - спермагенная клетка
На главной оси находятся мелкие чешуйки, называемые кроющими. В их пазухах развиваются крупные толстые чешуйки - семенные. На верхней стороне семенной чешуйки, у ее основания, расположены два семязачатка. Молодой семязачаток состоит из нуцеллуса и интегумента. Нуцеллус по существу и есть мегаспорангий. Он имеет яйцевидную форму и срастается с особым защитным покровом - интегументом. Лишь около вершины, обращенной к оси шишки, интегумект имеет отверстие, называемое микропиле, или пыльцевходом. Вначале нуцеллус состоит из однородных диплоидных клеток. Затем в средней части его обособляется одна более крупная археспориальная клетка, которая и является единственной материнской клеткой мегаспор. Она редукционно делится и образует четыре мегаспоры. В дальнейшем три из них дегенерируют, и только одна способна к развитию. Мегаспорангий никогда не вскрывается, так что мегаспора остается внутри него. Мегаспора многократно делится и формирует женский гаметофит, называемый здесь эндоспермом. Отметим, что у голосеменных эндосперм гаплоидный Из двух наружных клеток эндосперма, ориентированных к микропиле, образуются два архегония (рис.45,43, Г). Архегонии голосеменных значительно редуцированы по сравнению с папоротникообразными. Хорошо развита лишь яйцеклетка. Брюшная канальцевая клетка дегенерирует задолго до оплодотворения, шейка состоит из 8-12 клеток, шейковые канальцевые клетки не образуются. Таким образом, сосна по женской линии является внутреннезаростковым растением.

Рис. 45. Развитие женского гаметофита (эндосперма) сосны обыкновенной - Pinus
1 - археспориальная клетка, 2 - тетрада мегаспор, 3 - деление мегаспоры, 4 - женский гаметофж-(эндосперм), развившийся из мегаспоры
Пыльца из мужских шишек переносится на семязачатки и улавливается каплей густой жидкости, заполняющей пространство между нуцеллусом и интегументом и выступающей через микропиле (опыление). Подсыхая, она затягивает пыльцу внутрь семязачатка на нуцеллус (в пыльцевую камеру). После опыления микропиле зарастает. Чешуйки женской шишки смыкаются. Мужской гаметофит продолжает свое развитие на мегаспорангии. Экзина лопается, и вегетативная клетка, окруженная интиной, развивается в пыльцевую трубку (см. рис. 44, Ж), которая внедряется в ткань нуцеллуса и растет в направлении архегония. Антеридиальная клетка делится и образует две клетки: клетку-ножку и спермагенную клетку. Они переходят в пыльцевую трубку, которая и доставляет их к архегонию. Непосредственно перед оплодотворением из спермагенной клетки развиваются два спермия - мужские гаметы, лишенные жгутиков. Пыльцевая трубка представляет собой новообразование в связи с утратой подвижности гамет. Через шейку архегония она достигает яйцеклетки, приобретает повышенный тургор, кончик ее лопается, и содержимое выбрасывается в цитоплазму яйцеклетки. Вегетативное ядро разрушается. Один из спермиев сливается с ядром яйцеклетки (оплодотворение), а второй отмирает. От опыления до оплодотворения у сосны проходит около 13 месяцев. Из образовавшейся зиготы (диплоид) тотчас начинает развиваться зародыш. Развитие зародыша идет за счет запасных веществ эндосперма. На нуцеллус часто попадает несколько пылинок. Иногда могут быть оплодотворены оба архегония. Однако полноценный зародыш бывает только от одной зиготы. Сформировавшийся зародыш состоит из корешка, стебелька, нескольких семядолей (от 5 до 12) и почечки. Зародыш окружен эндоспермом, который потребляется при прорастании. Интегумент образует твердую кожуру. Так образуется семя. Семязачаток плотно прирастает к семенной чешуйке, из ткани которой развивается крыловидная пленка, способствующая распространению семян ветром. Созревание семян наступает осенью, на второй год после опыления. Шишки к этому времени достигают длины 4-6 см, чешуйки одревесневают, из зеленых становятся серыми. Следующей зимой шишки поникают, чешуйки расходятся, и семена высыпаются. Отделившись от материнского растения, семя может длительно находиться в состоянии покоя и лишь с наступлением благоприятных условий трогается в рост.
Очевидно, голосеменные произошли от каких-то групп высших споровых растений, существовавших в девонском периоде. Образование семени определило громадные преимущества голосеменных перед споровыми, что позволило им занять господствующее положение на суше уже в мезозойской эре. По