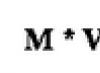Кора больших полушарий является высшим отделом центральной нервной системы, который в процессе филогенетического развития появляется позже всего и формируется в ходе индивидуального (онтогенетического) развития позже других отделов мозга. Кора представляет собой слой серого вещества толщиной 2-3 мм, содержащий в среднем около 14 млрд. (от 10 до 18 млрд.) нервных клеток, нервные волокна и межуточную ткань (нейроглию). На поперечном ее срезе по расположению нейрон
ов и их связей различают 6 горизонтальных слоев. Благодаря многочисленным извилинам и бороздам площадь поверхности коры достигает 0,2 м2. Непосредственно под корой находится белое вещество, состоящее из нервных волокон, которые передают возбуждение в кору и из нее, а также от одних участков коры другим.
Корковые нейрон
ы и их связи. Несмотря на огромное число нейрон
ов в коре, известно очень немного их разновидностей.
Основными типами их являются пирамидные и звездчатые нейрон
ы.
...
В афферентной функции коры и в процессах переключения возбуждения на соседние нейрон
ы основная роль принадлежит звездчатым нейрон
ам. Они составляют у человека более половины всех клеток коры. Эти клетки имеют короткие ветвящиеся аксон
ы, не выходящие за пределы серого вещества коры, и короткие ветвящиеся дендрит
ы. Звездчатые нейрон
ы участвуют в процессах восприятия раздражении и объединении деятельности различных пирамидных нейрон
ов.
Пирамидные нейрон ы осуществляют эфферентную функцию коры и внутрикорковые процессы взаимодействия между удаленными друг от друга нейрон ами. Они делятся на крупные пирамиды, от которых начинаются проекционные, или эфферентные, пути к подкорковым образованиям, и мелкие пирамиды, образующие ассоциативные пути к другим отделам коры. Наиболее крупные пирамидные клетки - гигантские пирамиды Беца - находятся в передней центральной извилине, в так называемой моторной зоне коры. Характерная особенность крупных пирамид - их вертикальная ориентация в толще коры. От тела клетки вертикально вверх к поверхности коры направлен наиболее толстый (верхушечный) дендрит , через который в клетку поступают различные афферентные влияния от других нейрон ов, а вертикально вниз отходит эфферентный отросток - аксон .
Многочисленность контактов (например, только на дендрит ах крупной пирамиды их насчитывают от 2 до 5 тыс.) обеспечивает возможность широкой регуляции деятельности пирамидных клеток со стороны множества других нейрон ов. Это позволяет координировать ответные реакции коры (в первую очередь ее моторную функцию) с разнообразными воздействиями из внешней среды и внутренней среды организма.
Для коры больших полушарий характерно обилие межнейрон ных связей. По мере развития мозга человека после его рождения увеличивается число межцентральных взаимосвязей, особенно интенсивно до 18 лет.
...
Первичные, вторичные и третичные поля коры
. Особенности строения и функционального значения отдельных участков коры позволяют выделить отдельные корковые поля.
Различают три основные группы полей в коре: первичные, вторичные и третичные поля.
Первичные поля связаны с органами чувств и органами движения на периферии, они раньше других созревают в онтогенез е, имеют наиболее крупные клетки. Это так называемые ядерные зоны анализаторов, по И. П. Павлову (например, поле болевой, температурной, тактильной и мышечно-суставной чувствительности в задней центральной извилине коры, зрительное поле в затылочной области, слуховое поле в височной области и двигательное поле в передней центральной извилине коры) (рис. 54). Эти поля осуществляют анализ отдельных раздражений, поступающих в кору от соответствующих рецептор ов. При разрушении первичных полей возникают так называемая корковая слепота, корковая глухота и т. п. Рядом расположены вторичные поля, или периферические зоны анализаторов, которые связаны с отдельными органами только через первичные поля. Они служат для обобщения и дальнейшей обработки поступающей информации. Отдельные ощущения синтезируются в них в комплексы, обусловливающие процессы восприятия. При поражении вторичных полей сохраняется способность видеть предметы, слышать звуки, но человек их не узнает, не помнит их значения. Первичные и вторичные поля имеются и у человека, и у животных.
Наиболее далеки от непосредственных связей с периферией третичные поля, или зоны перекрытия анализаторов. Эти поля есть только у человека. Они занимают почти половину территории коры и имеют обширные связи с другими отделами коры и с неспецифическими системами мозга. В этих полях преобладают наиболее мелкие и разнообразные клетки. Основным клеточным элементом здесь являются звездчатые нейрон ы. Третичные поля находятся в задней половине коры - на границах теменных, височных и затылочных ее областей и в передней половине - в передних частях лобных областей. В этих зонах оканчивается наибольшее число нервных волокон, соединяющих левое и правое полушария, поэтому роль их особенно велика в организации согласованной работы обоих полушарий. Третичные поля созревают у человека позже других корковых полей, они осуществляют наиболее сложные функции коры. Здесь происходят процессы высшего анализа и синтеза. В третичных полях на основе синтеза всех афферентных раздражении и с Учетом следов прежних раздражении вырабатываются цели и задачи поведения. Согласно им происходит программирование двигательной деятельности. Развитие третичных полей у человека связывают с функцией речи. Мышлени е (внутренняя речь) возможно только при совместной деятельности анализаторов, объединение информации от которых происходит в третичных полях.
При врожденном недоразвитии третичных полей человек не в состоянии овладеть речью (произносит лишь бессмысл енные звуки) и даже простейшими двигательными навыками (не может одеваться, пользоваться орудиями труда и т. п.).
Воспринимая и оценивая все сигналы из внутренней и внешней среды, кора больших полушарий осуществляет высшую регуляцию всех двигательных и эмоционально-вегетативных реакций.
Функции коры больших полушарии . Кора больших полушарий выполняет наиболее сложные функции организации приспособительного поведения организма во внешней среде. Это прежде всего функция высшего анализа и синтеза всех афферентных раздражении.
Афферентные сигналы поступают в кору по разным каналам, в разные ядерные зоны анализаторов (первичные поля), а затем синтезируются во вторичных и третичных полях, благодаря деятельности которых создается целостное восприятие внешнего мира. Этот синтез лежит в основе сложных психи ческих процессов восприятия, представления, мышлени я. Кора больших полушарий представляет собою орган, тесно связанный с возникновением у человека сознания и регуляцией его общественного поведения. Важной стороной деятельности коры больших полушарий является замыкательная функция - образование новых рефлексов и их систем (условные рефлексы, динамические стереотипы-см. главу XV).
Благодаря необычайно большой продолжительности сохранения в коре следов прежних раздражении (памяти) в ней накапливается огромный объем информации. Это имеет большое значение для сохранения индивидуального опыта, который используется по мере необходимости.
...
Экспериментально показано, что у высших представителей животного мира после полного оперативного удаления коры высшая нервная деятельность резко ухудшается. Они теряют способность тонко приспосабливаться к внешней среде и самостоятельно существовать в ней.
Современным ученым доподлинно известно, что благодаря функционированию головного мозга возможны такие способности, как осознание сигналов, которые получены из внешней среды, мыслительная деятельность, запоминание мышления.
Способность личности осознавать собственные отношения с другими людьми непосредственно связано с процессом возбуждения нейронных сетей. Причем речь идет именно о тех нейронных сетях, которые расположены в коре. Она представляет собой структурную основу сознания и интеллекта.
В данной статье рассмотрим, как устроена кора головного мозга, зоны коры головного мозга будут подробно описаны.
Неокортекс
Кора включает в себя около четырнадцати миллиардов нейронов. Именно благодаря им осуществляется функционирование основных зон. Подавляющая часть нейронов, до девяноста процентов, формирует неокортекс. Он является частью соматической НС и ее высшим интегративным отделом. Важнейшими функциями коры головного мозга выступают восприятие, переработка, интерпретация информации, которую человек получает при помощи всевозможных органов чувств.
Помимо этого, неокортекс управляет сложными движениями системы мышц человеческого тела. В нем расположены центры, принимающие участие в процессе речи, хранении памяти, абстрактном мышлении. Большая часть процессов, которые в нем происходят, формирует нейрофизическую основу человеческого сознания.
Из каких отделов еще состоит кора головного мозга? Зоны коры головного мозга рассмотрим ниже.
Палеокортекс
Является еще одним большим и важным отделом коры. В сравнении с неокортексом у палеокортекса более простая структура. Процессы, которые здесь протекают, редко отражаются в сознании. В этом отделе коры высшие вегетативные центры локализуются.
Связь коркового слоя с другими отделами мозга
Немаловажно рассмотреть связь, которая имеется между нижележащими отделами мозга и корой больших полушарий, например, с таламусом, мостом, средним мостом, базальными ядрами. Осуществляется эта связь при помощи крупных пучков волокон, которые внутреннюю капсулу формируют. Пучки волокон представлены широкими пластами, которые сложены из белого вещества. В них расположено огромное количество нервных волокон. Некоторая часть этих волокон обеспечивает передачу нервных сигналов к коре. Остальная часть пучков передает нервные импульсы к расположенным ниже нервным центрам.
Как устроена кора головного мозга? Зоны коры головного мозга будут представлены далее.
Строение коры
Самым большим отделом мозга является его кора. Причем зоны коры являются лишь одним типом частей, выделяемых в коре. Помимо этого кора разделена на два полушария - правое и левое. Между собой полушария соединены пучками белого вещества, формирующими мозолистое тело. Его функция - обеспечивать координацию деятельности обоих полушарий.
Классификация зон коры головного мозга по их расположению
Несмотря на то что кора имеет огромное количество складок, в общем расположение ее отдельных извилин и борозд постоянно. Главные их них являются ориентиром при выделении областей коры. К таким зонам (долям) относятся - затылочная, височная, лобная, теменная. Несмотря на то что они классифицируются по месту расположения, каждая из них имеет свои собственные специфические функции.
Слуховая зона коры головного мозга
К примеру, височная зона является центром, в котором расположен корковый отдел анализатора слуха. Если происходит повреждение этого отдела коры, может возникнуть глухота. Помимо этого в слуховой зоне расположен центр речи Вернике. Если повреждению подвергается он, то человек теряется способность к восприятию устной речи. Человек воспринимает ее как простой шум. Также в височной доле есть нейронные центры, которые относятся к вестибулярному аппарату. Если повреждаются они, нарушается чувство равновесия.

Речевые зоны коры головного мозга
В лобной доле коры сосредоточены речевые зоны. Речедвигательный центр расположен тоже здесь. Если происходит его повреждение в правом полушарии, то человек теряет способность изменять тембр и интонацию собственной речи, которая становится монотонной. Если же повреждение речевого центра произошло в левом полушарии, то пропадает артикуляция, способность к членораздельной речи и пению. Из чего еще состоит кора головного мозга? Зоны коры головного мозга имеют различные функции.
Зрительные зоны
В затылочной доле располагается зрительная зона, в которой находится центр, отвечающий на наше зрение как таковое. Восприятие окружающего мира происходит именно этой частью мозга, а не глазами. Именно затылочная зона коры ответственна за зрение, и ее повреждение может привести к частичной или полной потере зрения. Зрительная зона коры головного мозга рассмотрена. Что дальше?
Для теменной доли тоже характерны свои собственные специфические функции. Именно эта зона отвечает за способность анализировать информацию, которая касается тактильной, температурной и болевой чувствительности. Если происходит повреждение теменной области, рефлексы головного мозга нарушаются. Человек не может на ощупь распознавать предметы.
Двигательная зона
Поговорим о двигательной зоне отдельно. Следует отметить, что эта зона коры никак не соотносится с долями, рассмотренными выше. Она является частью коры, содержащей прямые связи с мотонейронами в спинном мозге. Такое название носят нейроны, непосредственно управляющие деятельностью мышц тела.

Основная двигательная зона коры больших полушарий располагается в извилине, которая называется прецентральной. Эта извилина представляет собой зеркальное отображение сенсорной зоны по многим аспектам. Между ними имеется контралатеральная иннервация. Если сказать иными совами, то иннервация направлена на мышцы, которые расположены на другой стороне тела. Исключение - лицевая область, для которой характерен контроль мышц двусторонний, расположенных на челюсти, нижней части лица.
Немного ниже основной двигательной зоны расположена дополнительная зона. Ученые полагают, что она имеет независимые функции, которые связаны с процессом вывода двигательных импульсов. Дополнительная двигательная зона также изучалась специалистами. Эксперименты, которые ставились над животными, показывают, что стимуляция этой зоны провоцирует возникновение двигательных реакций. Особенностью является то, что подобные реакции возникают даже в том случае, если основная двигательная зона была изолирована или разрушена полностью. Она также вовлечена в планирование движений и в мотивацию речи в полушарии, которое является доминантным. Ученые полагают, что при повреждении дополнительной двигательной может возникнуть динамическая афазия. Рефлексы головного мозга страдают.
Классификация по строению и функциям коры головного мозга
Физиологические эксперименты и клинические испытыния, которые проводились еще в конце девятнадцатого века, позволили установить границы между областями, на которые проецируются разные рецепторные поверхности. Среди них выделяют органы чувств, которые направлены на внешний мир (кожная чувствительность, слух, зрение), рецепторы, заложенные непосредствен в органах движения (двигательный или кинетический анализаторы).
Зоны коры, в которых располагаются разнообразные анализаторы, могут быть классифицированы по строению и функциям. Так, их выделяют три. К ним относятся: первичная, вторичная, третичная зоны коры головного мозга. Развитие эмбриона предполагает закладывание только первичных зон, характеризующихся простой цитоархитектоникой. Далее происходит развитие вторичных, третичные развиваются в самую последнюю очередь. Для третичных зон характерно самое сложное строение. Рассмотрим каждую из них немного подробнее.
Центральные поля
За долгие годы клинических исследований ученым удалось накопить значительный опыт. Наблюдения позволили установить, например, что повреждения различных полей, в составе корковых отделов разных анализаторов, могут отразиться далеко не равнозначно на общей клинической картине. Если рассматривать все эти поля, то среди них можно выделить одно, которое занимает центральное положение в ядерной зоне. Такое поле носит название центрального или первичного. Находится оно одновременно в зрительной зоне, в кинестетической, в слуховой. Повреждение первичного поля влечет за собой весьма серьезные последствия. Человек не может воспринимать и осуществлять самые тонкие дифференцировки раздражителей, влияющих на соответствующие анализаторы. Как еще классифицируются участки коры головного мозга?

Первичные зоны
В первичных зонах расположен комплекс нейронов, который наиболее предрасположен к обеспечению двусторонних связей между корковыми и подкорковыми зонами. Именно этот комплекс наиболее прямым и коротким путем соединяет кору больших полушарий с разнообразными органами чувств. В связи с этим данные зоны обладают способностью очень подробной идентификации раздражителей.
Важной общей чертой функциональной и структурной организации первичных областей является то, все они имеют четкую соматическую проекцию. Это означает, что отдельные периферические точки, например, кожные поверхности, сетчатка глаза, скелетная мускулатура, улитки внутреннего уха, имеют собственную проекцию в строго ограниченные, соответствующие точки, которые находятся в первичных зонах коры соответствующих анализаторов. В связи с этим им было дано название проекционных зон коры головного мозга.
Вторичные зоны
По-другому эти зоны называются периферическими. Такое название дано им совсем не случайно. Они находятся в периферических отделах участков коры. От центральных (первичных) вторичные зоны отличаются нейронной организацией, физиологическими проявлениями и особенностями архитектоники.
Попробуем разобраться, какие эффекты возникают, если на вторичные зоны воздействует электрический раздражитель или происходит их повреждение. Главным образом возникающие эффекты касаются наиболее сложных видов процессов в психике. В том случае, если происходит повреждение вторичных зон, то элементарные ощущения остаются в относительной сохранности. В основном наблюдаются нарушения в способности правильного отражения взаимных соотношений и целых комплексов элементов, из которых состоят различные объекты, которые мы воспринимаем. К примеру, если повреждению подверглись вторичные зоны зрительной и слуховой коры, то можно наблюдать возникновение слуховых и зрительных галлюцинаций, которые разворачиваются в определенной временной и пространственной последовательности.

Вторичные области имеют значительную важность в реализации взаимных связей раздражителей, которые выделяются при помощи первичных зон коры. Помимо этого, значительную роль они играют в интеграции функций, которые осуществляют ядерные поля разных анализаторов в результате объединения в сложные комплексы рецепций.
Таким образом, вторичные зоны представляют особую важность для реализации психических процессов в более сложных формах, которые требуют координации и которые связаны с подробным анализом соотношений между предметными раздражителями. В ходе этого процесса устанавливаются специфические связи, которые носят название ассоциативных. Афферентные импульсы, поступающие в кору от рецепторов разных внешних органов чувств, достигают вторичных полей посредством множества дополнительных переключений в ассоциативном ядре таламуса, который также называется зрительным бугром. Афферентные импульсы, следующие в первичные зоны, в отличие от импульсов, следуют во вторичные зоны, достигают их путем, который короче. Он реализован посредством реле-ядра, в зрительном бугре.
Мы разобрались, за что отвечает кора головного мозга.
Что такое таламус?
От таламических ядер к каждой доле мозговых полушарий подходят волокна. Таламус является зрительным бугром, расположенным в центральной части переднего отдела мозга, состоит из большого количества ядер, каждое из которых осуществляет передачу импульса в определенные участки коры.
Все сигналы, которые поступают к коре (исключение составляют только обонятельные), проходят через релейные и интегративные ядра зрительного бугра. От ядер таламуса волокна направляются к сенсорным зонам. Вкусовая и соматосенсорная зоны расположены в теменной доле, слуховая сенсорная зона - в височной доле, зрительная - в затылочной.

Импульсы к ним поступают, соответственно, от вентро-базальных комплексов, медиальных и латеральных ядер. Моторные зоны связаны с вентеральным и вентролатеральным ядрами таламуса.
Десинхронизация ЭЭГ
Что произойдет, если на человека, находящегося в состоянии полного покоя, подействует очень сильный раздражитель? Естественно, что человек полностью сконцентрируется на данном раздражителе. Переход умственной деятельности, который осуществляется от состояния покоя к состоянию активности, отражается на ЭЭГ бета-ритмом, который замещает альфа-ритм. Колебания становятся более частыми. Такой переход называют десинхронизацией ЭЭГ, появляется он в результате поступления сенсорного возбуждения в кору от неспецифических ядер, расположенных в таламусе.
Активирующая ретикулярная система
Диффузную нервную сесть составляют неспецифические ядра. Находится эта система в медиальных отделах таламуса. Он является передним отделом активирующей ретикулярной системы, регулирующей возбудимость коры. Разнообразные сенсорные сигналы способны активировать данную систему. Сенсорные сигналы могут быть как зрительными, так и обонятельными, соматосенсорными, вестибулярными, слуховыми. Активизирующая ретикулярная система представляет собой канал, который передает к поверхностному слою коры данные сигналов через расположенные в таламусе неспецифические ядра. Возбуждение АРС необходимо для того, чтобы человек был способен поддерживать состояние бодрствования. Если в данной системе возникают нарушения, то могут наблюдаться коматозные сноподобные состояния.
Третичные зоны
Между анализаторами коры головного мозга имеются функциональные отношения, которые имеют еще более сложную структуру, чем та, что была описана выше. В процессе роста происходит взаимное перекрытие полей анализаторов. Такие зоны перекрытия, которые образуются у концов анализаторов, носят название третичных зон. Они являются самыми сложными типами объединения деятельности слухового, зрительного, кожно-кинестетического анализаторов. Расположены третичные зоны за границами собственных зон анализаторов. В связи с этим повреждение их не оказывает выраженного эффекта.

Третичные зоны представляют собой особые корковые области, в которых собраны рассеянные элементы разных анализаторов. Они занимают весьма обширную территорию, которая разделена на области.
Верхняя теменная область интегрирует движения всего тела с анализатором зрительным, формирует схему тел. Нижняя теменная область объединяет обобщенные формы сигнализации, которые связаны с дифференцированными предметными и речевыми действиями.
Не менее важной является височно-теменно-затылочная область. Отвечает она за усложненные интеграции слухового и зрительного анализаторов с устной и письменной речью.
Стоит отметить, что по сравнению с двумя первыми зонами, для третичных характерны наиболее сложные цепи взаимодействия.
Если опираться на весь изложенный выше материал, то можно сделать вывод о том, что первичные, вторичные, третичные зоны коры у человека носят высокую специализацию. Отдельно стоит подчеркнуть тот факт, что все три корковые зоны, которые мы рассматривали, в нормально функционирующем мозге совместно с системами связей и образованиями подкоркового расположения функционируют как единое дифференцированное целое.
Мы подробно рассмотрели зоны и отделы коры головного мозга.
Тема нашей лекции « Функции коры больших полушарий».
Головной мозг находится в полости мозгового черепа. Имеет выпуклую верхнелатеральную и нижнюю поверхности, а также уплощенную поверхность - основание головного мозга.
Большой мозг состоит из двух полушарий - правого и левого, которые связаны комиссурой - мозолистым телом. Правое и левое полушария делятся с помощью продольной щели. Под комиссурой находится свод, представляющий собой два изогнутых волокнистых тяжа, которые в средней части соединены между собой, а спереди и сзади расходятся, образуя столбы и ножки свода. Спереди от столбов свода находится передняя спайка. Между мозолистым телом и сводом натянута тонкая вертикальная пластинка мозговой ткани - прозрачная перегородка.
Полушария имеют верхнелатеральную, медиальную и нижнюю поверхности. Верхнелатеральная - выпуклая, медиальная - плоская, обращенная к такой же поверхности другого полушария, и нижняя - неправильной формы. На трех поверхностях располагаются глубокие и мелкие борозды, и между ними извилины. Борозды - углубления между извилинами. Извилины - возвышения мозгового вещества.
Поверхности полушарий большого мозга отделены друг от друга краями - верхним, нижнелатеральным и нижневертикальным. В пространстве между двумя полушариями входит большой серповидный отросток, представляющий собой тонкую пластинку твердой оболочки, которая проникает в продольную щель большого мозга, не достигая мозолистого тела, и отделяет друг от друга правое и левое полушария.
Наиболее выступающие участки полушарий получили название полюсов: лобного, затылочного и височного. Рельеф поверхностей полушарий большого мозга очень сложен и связи с наличием более или менее глубоких борозд большого мозга и расположенных между ними валикообразных возвышений - извилин. Глубина, протяженность некоторых борозд и извилин, их форма и направление очень изменчивы.
Каждое полушарие делится на доли - лобную, теменную, затылочную, височную , островковую. Центральная борозда отделяет лобную долю от теменной, латеральная борозда - височную долю от лобной и теменной, теменно-затылочная борозда разделяет теменную и затылочную доли.
Кора больших полушарий состоит горизонтальных слоев, расположенных в направлении с поверхности в глубь.
I. Молекулярный слой имеет очень мало клеток, но большое количество ветвящихся дендритов пирамидных клеток, формирующих сплетение, расположенное параллельно поверхности. На этих дендритах образуют синапсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер таламуса.
II. Наружный зернистый слой составлен в основном звездчатыми и частично малыми пирамидными клетками. Волокна клеток этого слоя расположены преимущественно вдоль поверхности коры, образуя кортикокортикальные связи.
III. Наружный пирамидный слой состоит преимущественно из пирамидных клеток средней величины. Аксоны этих клеток, как и зернистые клетки II слоя, образуют кортикокортикальные ассоциативные связи.
IV. Внутренний зернистый слой по характеру клеток и расположению их волокон аналогичен наружному зернистому слою. На нейронах этого слоя образуют синаптические окончания афферентные волокна, идущие от нейронов специфических ядер таламуса и, следовательно, от рецепторов сенсорных систем.
V. Внутренний пирамидный слой образован средними и крупными пирамидными клетками, причем гигантские пирамидные клетки Беца расположены в двигательной коре. Аксоны этих клеток образуют эфферентные кортикоспинальные и кортикобульбарный двигательные пути.
VI. Слой полиморфных клеток образован преимущественно веретенообразными клетками, аксоны которых образуют кортикоталамические пути.
В первом и четвертом слоях происходят восприятие и обработка поступающих в кору сигналов. Нейроны второго и третьего слоев осуществляют кортикокортикальные ассоциативные связи. Покидающие кору эфферентные пути формируются преимущественно в пятом и шестом слоях.
Более детально деление коры на различные поля проведено на основе формы и расположения нейронов Бродманом, который выделил 11 областей, включающих в себя 52 поля, многие из которых характеризуются функциональными и нейрохимическими особенностями. По Бродману лобная область включает следующие поля: 8, 9, 10, 11, 12, 44, 45, 46, 47. В прецентральную область входят 4 и 6 поле, в постцентральную область входят 1, 2, 3 и 43 поле. Теменная область включает в себя поля 5, 7, 39, 40, а затылочная область 17 18 19. Височная область состоит из очень большого количества полей.
В коре выделяют сенсорные, ассоциативные и двигательные зоны, исходя из расположения нейронов:
Проблема локализации функций в коре больших полушарий имеет три концепции:
Принцип узкой локализации - все функции помещены в одну, отдельно взятую структуру.
Концепция эквипотенциала - различные корковые структуры функционально равноценны.
Принцип многофункциональности корковых полей.
Свойство мультифункциональности позволяет данной структуре включаться в обеспечение различных форм деятельности, реализуя при этом основную, генетически присущую ей функцию. Степень мультифункциональности различных корковых структур неодинакова: например, в полях ассоциативной коры она выше, чем в первичных сенсорных полях, а в корковых структурах выше, чем в стволовых. В основе мультифункциональности лежит многоканальность поступления в кору мозга афферентного возбуждения, перекрытие афферентных возбуждений, особенно на таламическом и корковым уровнях, модулирующее влияние различных структур (неспецифического таламуса, базальных ганглиев) на корковые функции, взаимодействие корково-подкорковых и межкорковых путей проведения возбуждения.
Одним из наиболее крупных вариантов функционального разделения новой коры головного мозга является выделение в ней сенсорной, ассоциативной и двигательной областей.
Сенсорные области коры - это зоны, в которые проецируются сенсорные раздражители. Сенсорные области коры иначе называют: проекционной корой или корковыми отделами анализаторов. Они расположены преимущественно в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от специфических сенсорных ядер таламуса. Сенсорная кора имеет хорошо выраженные вторые и четвертые слои и называется гранулярной.
Зоны сенсорной коры, раздражение или разрушение которых вызывает четкие и постоянные изменения чувствительности организма, называются первичными сенсорными областями. Они состоят преимущественно из мономодальных нейронов и формируют ощущения одного качества. В первичных сенсорных зонах обычно имеется четкое пространственное представительство частей тела, их рецепторных полей. Вокруг первичных сенсорных зон находятся менее локализованные вторичные сенсорные зоны, полимодальные нейроны которых отвечают на действие нескольких раздражителей.
Важнейшей сенсорной областью является теменная кора постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной поверхности полушарий, которую обозначают как первичная соматосенсорная область. Здесь имеется проекция кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных рецепторов, интероцептивной чувствительности и чувствительности опорно-двигательного аппарата от мышечных, суставных и сухожильных рецепторов. Проекция участков тела в этой области характеризуется тем, что проекция головы и верхних отделов туловища расположена в нижнелатеральных участках постцентральной извилины, проекция нижней половины туловища и ног - в верхнемедиальных зонах извилины, проекция нижней части голени и стоп - в коре парацентральной дольки на медиальной поверхности полушарий. При этом проекция наиболее чувствительных участков (язык, губы, гортань, пальцы рук) имеет относительно большие зоны по сравнению с другими частями тела. Предполагается, что в зоне тактильной чувствительности языка расположена и проекция вкусовой чувствительности.
Вторичная соматосенсорная область меньших размеров расположена на верхней стенке боковой борозды, на границе ее пересечения с центральной бороздой. Функции вторичной соматосенсорной области изучены плохо. Известно, что локализация поверхности тела в ней менее четкая, импульсация сюда поступает как от противоположной стороны тела, так и от «своей» стороны, предполагают ее участие в сенсорной и моторной координации двух сторон тела.
Другой первичной сенсорной зоной является слуховая кора, которая расположена в глубине латеральной борозды. В этой зоне в ответ на раздражение слуховых рецепторов кортиева органа формируются звуковые ощущения, изменяющиеся по громкости, тону и другим качествам. Здесь имеет четкая топическая проекция: в разных участках коры представлены различные участки кортиева органа. К проекционной коре височной доли относится также центр вестибулярного анализатора в верхней и средней височных извилинах. Обработанная сенсорная информация используется для формирования «схемы тела» и регуляции функций мозжечка (височно-мостомозжечковый путь).
Еще одна первичная проекционная область новой коры расположена в затылочной коре - первичная зрительная область. Здесь имеет топическое представительство рецепторов сетчатки, и каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна имеет большую зону представительства. В связи с неполным перекрестом зрительных путей в зрительную область каждого полушария проецируются одноименные половины сетчатки. Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения.
Раздражение коры 17-го поля приводит к возникновению световых ощущений. Около поля 17 расположена кора вторичной зрительной области. Нейроны этих зон полимодальны и отвечают не только на световые, но и на тактильные, слуховые раздражители. В данной зрительной области происходит синтез различных видов чувствительности и возникают более сложные зрительные образы и их опознавание. Раздражение этих полей вызывает зрительные галлюцинации, навязчивые ощущения, движения глаз.
Основная часть информации об окружающей среде и внутренней среда организма, поступившая в сенсорную кору, передается для дальнейшей ее обработки в ассоциативную кору.
Ассоциативные области коры включают участки новой коры, расположенные рядом с сенсорными и двигательными зонами, но не выполняющие непосредственно чувствительных и двигательных функций. Границы этих областей обозначены не достаточно четко, неопределенность преимущественно связана со вторичными проекционными зонами, функциональные свойства которых являются переходными между свойствами первичных проекционных и ассоциативных зон. У человека ассоциативная кора составляет 70% неокортекса.
Основной физиологической особенностью нейронов ассоциативной коры является полимодальность: они отвечают на несколько раздражителей с почти одинаковой силой. Полимодальность (полисенсорность) нейронов ассоциативной коры создается за счет, во-первых, наличия кортикокортикальных связей с разными проекционными зонами, во-вторых, за счет главного афферентного входа от ассоциативных ядер таламуса, в которых уже произошла сложная обработка информации от различных чувствительных путей. В результате этого ассоциативная кора представляет собой мощный аппарат конвергенции различных сенсорных возбуждений, позволяющих произвести сложную обработку информации о внешней и внутренней среде организма и использовать ее для осуществления высших психофизиологических функций. В ассоциативной коре выделяют три ассоциативные системы мозга: таламотеменную, таламолобную и таламовисочную.
Таламотеменная система представлена ассоциативными зонами теменной коры, получающими основные афферентные входы от задней группы ассоциативных ядер таламуса. Теменная ассоциативная кора имеет эфферентные выходы на ядра таламуса и гипоталамуса, моторную кору и ядра экстрапирамидной системы.
Основными функциями таламотеменной системы являются гнозис, формирование «схемы тела» и праксис.
Под гнозисом понимают функцию различных видов узнавания: формы, величины, значения предметов, понимание речи, познание процессов, закономерностей. К гностическим функциям относится оценка пространственных отношений. В теменной коре выделяют центр стереогнозиса, расположенный сзади от средних отделов постцентральной извилины (поля 7, 40, частично 39) и обеспечивающий способность узнавания предметов на ощупь. Вариантом гностической функции является формирование в сознании трехмерной модели тела, центр которой расположен в поле 7 теменной коры. Под праксисом понимают целенаправленное действие, центр его находится в надкраевой извилине (поля 39 и 40 доминантного полушария). Этот центр обеспечивает хранение и реализацию программы двигательных автоматизированных актов.
Таламолобная система представлена ассоциативными зонами лобной коры, имеющими основной афферентный вход от ассоциативного медиодорсального ядра таламуса. Главной функцией лобной ассоциативной коры является формирование программ целенаправленного поведения, особенно в новой для человека обстановке. Реализация этой общей функции основывается на других функциях таламолобной системы: 1) формирование доминирующей мотивации обеспечивающей направление поведения человека. Эта функция основана на тесных двусторонних связях лоьной коры с лимбической системой и ролью последней в регуляции высших эмоций человека, связанных с его социальной деятельностью и творчеством; 2) обеспечение вероятностного прогнозирования, что выражается изменением поведения в ответ на изменения обстановки окружающей среды и доминирующей мотивации; 3) самоконтроль действий путем постоянного сравнения результата действия с исходными намерениями, что связано с созданием аппарата предвидения (акцептора результата действия).
При повреждении префронтальной лобной коры, где пересекаются связи между лобной долей и таламусом, человек становится грубым, нетактичным, ненадежным, у него появляется тенденция к повторению каких-либо двигательных актов, хотя обстановка уже изменилась и надо выполнять другие действия.
Таламовисочная система изучена не достаточно. Но если говорить о височной коре, то надо отметить, что некоторые ассоциативные центры, например стереогнозиса и праксиса, включают в себя и участки височной коры. В височной коре расположен слуховой центр речи, находящийся в задних отделах верхней височной извилины. Этот центр обеспечивает речевой гнозис - распознавание и хранение устной речи, как собственной, так и чужой. В средней части верхней височной извилины находится центр распознавания музыкальных звуков и их сочетаний. На границе височной, теменной и затылочной долей находится центр чтения письменной речи, обеспечивающий распознавание и хранение образов письменной речи.
В двигательной коре выделяют первичную и вторичную моторные области.
В первичной моторной коре расположены нейроны, иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая топографическая проекция мышц тела. При этом проекции мышц нижних конечностей и туловища расположены в верхних участках прецентральной извилины и занимают сравнительно небольшую площадь, а проекция мышц верхних конечностей, лица и языка расположены в нижних участках извилины и занимают большую. Основной закономерностью топографического представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков двигательной коры. Двигательные реакции на раздражение первичной моторной коры осуществляются с минимальным порогом (высокая возбудимость), и представлены элементарными сокращениями мышц противоположной стороны тела (для мышц головы сокращение может быть билатеральным). При поражении этой области коры утрачивается способность к тонким координированным движениям рук, особенно пальцев.
Вторичная двигательная кора расположена на латеральной поверхности полушарий, впереди прецентральной извилины. Она осуществляет высшие двигательные функции, связанные с планированием и координацией произвольных движений. Кора поля 6 получает основную часть эфферентной импульсации базальных ядер и мозжечка и участвует в перекодировании информации о программе сложных движений. Раздражение коры поля 6 вызывает более сложные координированные движения, например, поворот головы, глаз и туловища в противоположную сторону, содружественные сокращения мышц-сгибателей или мышц-разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры, связанные с социальными функциями человека: центр письменной речи в заднем отделе средней лобной извилины, центр моторной течи Брока в заднем отделе нижней лобной извилины, обеспечивающий речевой праксис, а также музыкальный моторный центр, определяющий тональность речи, способность петь.
В моторной коре лучше, чем в других зонах коры, выражен слой, содержащий гигантские пирамидные клетки Беца. Нейроны двигательной коры получают афферентные входы через таламус от мышечных, суставных и кожных рецепторов, а также от базальных ядер и мозжечка. Основной эфферентный выход двигательной коры на стволовые и спинальные моторные центры формируют пирамидные клетки пятого слоя. Пирамидные и сопряженные с ними вставочные нейроны расположены вертикально по отношению к поверхности коры и образуют нейронные двигательные колонки. Пирамидные нейроны двигательной колонки могут возбуждать или тормозить мотонейроны стволовых и спинальных центров. Соседние колонки в функциональном плане перекрываются, а пирамидные нейроны, регулирующие деятельность одной мышцы, расположены обычно не в одной, а в нескольких колонках.
Основные эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные пути, которые начинаются от гигантских пирамидных клеток Беца и менее крупных пирамидных клеток пятого слоя коры прецентральной извилины (60% волокон), премоторной коры (20% волокон) и постцентральной извилины (20% волокон). Крупные пирамидные клетки имеют быстропроводящие аксоны и фоновую импульсную активность около 5 Гц, которая при движении увеличивается до 20-30 Гц. Эти клетки иннервируют крупные (высокопороговые) ?-мотонейроны в двигательных центрах ствола и спинного мозга, регулирующих физические движения. От мелких пирамидных клеток отходят тонкие медленнопроводящие миелиновые аксоны. Эти клетки имеют фоновую активность около 15 Гц, которая во время движения увеличивается или уменьшается. Они иннервируют мелкие (низкопороговые) ?-мотонейроны в стволовых и спинальных двигательных центрах, регулирующие тонус мышц.
Пирамидные пути состоят из 1 млн волокон кортикоспинального пути, которые начинаются от коры верхней и средней трети прецентральной извилины, и 20 млн волокон кортикобульбарного пути, который начинается от коры нижней трети прецентральной извилины.
Волокна пирамидного пути оканчиваются на альфа-мотонейронах двигательных ядер третьего - седьмого и девятого - двенадцатого черепных нервов (кортикобульбарный путь) или на спинальных двигательных центрах (кортикоспинальный путь).
Через двигательную кору и пирамидные пути осуществляются произвольные простые движения и сложные целенаправленные двигательные программы, например, профессиональные навыки, формирование которых начинается в базальных ганглиях и мозжечке и заканчивается во вторичной моторной коре.
Большинство волокон пирамидных путей осуществляют перекрест, однако небольшая часть волокон остается неперекрещенными, что способствует компенсации нарушенных функций движения при односторонних поражениях. Через пирамидные пути осуществляет свои функции и премоторная кора: двигательные навыки письма, поворот головы, глаз и туловища в противоположную сторону, а также речь. В регуляции письма и особенно устной речи имеется выраженная асимметрия больших полушарий мозга: у 95% правшей и 70% левшей устная речь контролируется левым полушарием.
К корковым экстрапирамидным путям относят кортикорубральные и кортикоретикулярные пути, начинающиеся приблизительно от тех зон, которые дают начало пирамидным путям. Волокна кортикорубрального пути оканчиваются на нейронах красных ядер среднего мозга, от которых далее идут руброспинальные пути.
Волокна кортикоретикулярных путей оканчиваются на нейронах медиальных ядер ретикулярной формации моста (от них идут медиальные ретикулоспинальные пути) и на нейронах ретикулярных гигантоклеточных ядер продолговатого мозга, от которых начинаются латеральные ретикулоспинальные пути.
Через эти пути осуществляется регуляция тонуса и позы, которые обеспечивают точные целенаправленные движения. Корковые экстрапирамидные пути являются компонентом экстрапирамидной системы головного мозга, к которой относятся мозжечок, базальные ганглии, моторные центры ствола. Экстрапирамидная система осуществляет регуляцию тонуса, позы равновесия, выполнение заученных двигательных актов, таких как ходьба, бег, речь, письмо. Поскольку кортикопирамидные пути отдают свои многочисленные коллатерали структурам экстрапирамидной системе, то обе системы работают в функциональном единстве.
Оценивая в общем плане роль различных структур головного и спинного мозга в регуляции сложных направленных движений, можно отметить, что побуждение (мотивация) к движению создается в лимбической системе, замысел движения - в ассоциативной коре больших полушарий, программы движений - в базальных ганглиях, мозжечке и премоторной коре, а выполнение сложных движений происходит через двигательную кору, моторные центры ствола и спинного мозга.
Межполушарные взаимоотношения у человека проявляются в двух формах - функциональной асимметрии больших полушарий и совместной их деятельности.
Межполушарная асимметрия как одна из важных особенностей функционирования высших отделов мозга в основном определяется двумя моментами: 1) асимметричной локализацией нервного аппарата второй сигнальной системы и 2) доминированием правой руки как мощного средства адаптивного поведения человека. Этим и объясняется, что первые представления о функциональной роли межполушарной асимметрии возникли лишь тогда, когда удалось установить локализацию нервных центров речи (моторного — центра Брока и сенсорного — центра Вернике в левом полушарии).
Перекрестная проекция видов сенсорной чувствительности и нисходящих пирамидных путей — регуляторов моторной сферы организма — в сочетании с левосторонней локализацией центра устной и письменной речи определяет доминирующую роль левого полушария в поведении человека, управляемого корой больших полушарий.
Полученные экспериментальные данные подтверждают представление о доминирующей роли левого полушария мозга в реализации функций второй сигнальной системы, в мыслительных операциях, в творческой деятельности с преобладанием форм абстрактного мышления. В общем виде можно считать, что люди с левополушарным доминированием относятся к мыслительному типу, а с правополушарным доминированием — к художественному.
Функциональная асимметрия полушарий является важнейшим психофизиологическим свойством головного мозга человека. Выделяют психическую, сенсорную и моторную межполушарную функциональную асимметрии мозга.
При исследовании психофизиологических функций было показано, что в речи словесный информационный канал контролируется левым полушарием, а несловесный канал (голос, интонация) - правым.
Абстрактное мышление и сознание связаны, преимущественно, с левым полушарием. При выработке условного рефлекса в начальной фазе доминирует правое полушарие, а во время упрочения рефлекса - левое.
Правое полушарие осуществляет обработку информации одновременно, синтетически, по принципу дедукции, лучше воспринимаются пространственные и относительные признаки предмета. Левое полушарие производит обработку информации последовательно, аналитически, по принципу индукции, лучше воспринимает абсолютные признаки предмета и временные отношения.
В эмоциональной сфере правое полушарие обуславливает преимущественно отрицательные эмоции, контролирует проявления сильных эмоций, в целом оно более «эмоционально». Левое полушарие обуславливает в основном положительные эмоции, контролирует проявление более слабых эмоций.
В сенсорной сфере роль правого и левого полушарий лучше всего проявляется при зрительном восприятии. Правое полушарие воспринимает зрительный образ целостно, сразу во всех подробностях, легче решает задачу различения предметов и опознания визуальных образов предметов, которое трудно описать словами, создает предпосылки конкретно-чувственного мышления. Левое полушарие оценивает зрительный образ расчленено, аналитически, при этом каждый признак анализируется раздельно. Легче опознаются знакомые предметы и решаются задачи сходства предметов, зрительные образы лишены конкретных подробностей и имеют высокую степень абстракции; создаются предпосылки логического мышления.
Моторная асимметрия выражается, прежде всего, в право-леворукости, которая контролируется моторной корой противоположного полушария. Асимметрия других групп мышц имеет индивидуальный, а не видовой характер.
Парность в деятельности больших полушарий обеспечивается наличием комиссуральной системы (мозолистого тела, передней и задней, гиппокампальной и хабенулярной комиссур, межталамического сращения), которые анатомически соединяют два полушария головного мозга. Иначе говоря, оба полушария связаны не только горизонтальными связями, но и вертикальными.
Основные факты, полученные с помощью электрофизиологических методик, показали, что возбуждение из участка раздражения одного полушария передается через комиссуральную систему не только в симметричный участок другого полушария, но и в несимметричные участки коры. Исследование метода условных рефлексов показало, в процессе выработки рефлекса происходит «перенос» временной связи в другое полушарие. Элементарные же формы взаимодействия двух полушарий могут осуществляться через четверохолмие и ретикулярную формацию ствола.
У человека, как и у многих животных, большинство органов парные: две руки, две ноги, два глаза, два уха, две почки, два полушария мозга. Парность органов не означает их одинаковое функционирование. Мы знаем, какая рука у нас ведущая - выполняет наиболее сложные, тонкие операции. У большинства людей - это правая рука. Мы едим, шьем, пишем, рисуем правой рукой. Среди людей - правшей, использующих для точных действий правую руку, 90%, тогда как левши составляют в среднем 10%.
Левши всех рас и культур в прошлом и настоящем находились в меньшинстве среди праворукого окружения.
При изучении вопроса о происхождении левшества выделились три основных направления: «генетическое», «культурное» и «патологическое».
В настоящее время наибольшее распространение получили две генетические модели. Согласно одной асимметрия мозга определяется присутствием одного гена, который был назван ею фактором «правого сдвига». Если данный фактор имеется у индивидуума, последний предрасположен быть правшой. Если фактор отсутствует, человек может быть либо левшой, либо правшой в зависимости от случайных обстоятельств. При этом большое значение придается повреждениям мозга в пренатальном и раннем постнатальном периоде, которые могут повлиять на фенотипическую реализацию фактора «правого сдвига».
Более сложная модель была предложена Леви и Нагилаки (1972). Эти ученые предполагают, что рукость является функцией двух генов. Один ген с двумя аллелями определяет полушарие, которое будет контролировать речь и ведущую руку.
На вероятность не генетической, а цитоплазматической закодированности асимметрии указывает Морган (1978), выдвигая концепцию, согласно которой и мозговая латерализация, и мануальное предпочтение рассматриваются в широком общебиологическом аспекте. Предполагается, что развитие мозга находится под влиянием лево-правого градиента, а это приводит к более раннему и быстрому созреванию в онтогенезе левого полушария, которое при этом оказывает тормозящее влияние на правое - в результате возникает доминирование левого полушария по речи и праворукость.
С «генетическим» направлением непосредственно сочетаются исследования, связанные с выявлением анатомических, физиологических и морфологических стигматов, свойственных правшам и левшам. Показано, что у правшей сильвиева борозда справа расположена выше левой, в то время, как у 71% левшей правая и левая борозды примерно симметричны.
У правшей отмечается больший диаметр внутренней сонной артерии слева и выше давление в ней, чем в правой, а у левшей - обратная картина.
Аналогичная диссоциация выявляется у правшей и левшей при изучении средней мозговой артерии. Гипотеза Гершвинда и Галабурды также предполагает эндокринное влияние на формирование различий в строении мозга мужчин и женщин. Известна теория Превика, согласно которой церебральная латерализация у человека формируется при асимметричном пренатальном развитии системы внутреннего уха и лабиринта.
Существует и генетико-культурная гипотеза функциональной асимметрии. Английский ученый из Кембриджа Лэлэнд и его коллеги считают, что левшество является в равной степени генетически и культурологически обусловленным.
Альтернативными «генетическим» представляются гипотезы возникновения межполушарной асимметрии, базирующиеся на признании детерминирующей роли культурных условий в формировании рукости. «Культурно-социальные» концепции рассматривают правшество-левшество как следствие социального воспитания, опыта, условий жизни.
Наряду с представленными выше теориями, широко распространены представления о патологическом происхождении левшества. Крайней точки зрения придерживается Бэкан (1973), который утверждает, что любое проявление леворукости есть следствие родовой травмы. По мнению Чуприкова (1975), изменение моторного доминирования является одним из объективных доказательств врожденной энцефалопатии. В подтверждение приводятся факты увеличения левшей среди близнецов, особенности пренатального развития которых предполагают риск внутриутробной гипоксии мозга. В пользу этого подхода говорят и результаты проб Вада, согласно которым повреждение левого полушария на ранних этапах онтогенеза может привести к смене ведущей руки и доминантного по речи полушария.
Изучение вопроса о происхождении латеральности продолжается. Обилие фактов, подчас противоречащих друг другу, показывает, что каждая из теорий функциональной межполушарной асимметрии мозга требует дальнейшего обоснования. Вместе с тем очевидно, что основополагающие принципы вышеперечисленных подходов составляют базу для будущего системного исследования, необходимость которого вытекает из множества проблем и вопросов, оставшихся открытыми.
Как устроен наш мозг? Сколько нейронов в нем и каковы функции неокортекса? Современные ученые скрупулезно исследуют особенности нашего мозга и открывают все больше интересных подробностей.
Благодаря развитию высших нервных центров человек определяет себя и свое место в социуме, сознательно контролирует свое поведение и способен к адаптации в новой среде. Все эти преимущества связаны с функциями больших полушарий, которые мы рассмотрим.
Особенности мозга человека
Мозг человеческого вида весит приблизительно 1 кг 200 грамм - это средние показатели. Он состоит из 5 основных частей: это конечный, промежуточный, средний, задний и продолговатый мозг.

Большие борозды (углубления) разделяют 4 основные части лобную долю от теменной; а теменную - от затылочной; примыкает к трем другим. Последняя, пятая доля - островковая, которая находится в глубине латеральной ямки. Гармоничное взаимодействие всех нейронов обеспечивает рост и развитие нашей индивидуальности, наш характер и способности.
Можно выделить отдельную функцию больших полушарий - непрекращающееся развитие. Мозг человека все время развивается. Все, что индивидуум читает, видит, воспринимает, он буквально впитывает в себя. Особенно важна новая информация для детей до 2 лет, в это время их нейроны активно выстраивают связи на будущее.
Большие полушария. Строение и функции
В коре имеется от 14 до 17 млрд нейронов; а связей между клетками во много раз больше. Нейроны соединены синапсами. А помогают активировать связи различные нейромедиаторы - химические вещества, которые активируют рядом находящийся синапс.
Полушария мозга имеют особую структуру. Благодаря складкам, состоящим из борозд и извилин, площадь коры значительно увеличивается. По некоторым данным, общая площадь коры у среднестатистического человека - 2200 кв. см.

Под корой находится подкорка, или белое вещество мозга. Полушария между собой соединены мозолистым телом. А еще глубже находятся желудочки мозга - заполненные спинномозговой жидкостью пространства.
Кора состоит из слоев нервных нервных клеток, которые чередуются со слоями их ответвлений - аксонов. Всего насчитывается 6 слоев:
- молекулярный слой;
- наружный зернистый;
- наружный пирамидный - содержит преимущественно пирамидные нейроны;
- внутренний зернистый;
- внутренний пирамидный;
- слой веретеновидных нейронов.

Веретеновидные нейроны постепенно переходят в мозга. В коре происходят сознательные действия, формируется речь. В нижних глубинных частях под корой расположены центры бессознательных рефлексов и контроль внутренних органов и систем органов.
Зоны мозга
Чтобы понять функции больших полушарий головного мозга, нужно сначала разобрать их структуру. Полушария разделены условно на несколько центров, в которых проходят определенные психические и физиологические процессы. Эти центры не являются какими-то отдельными структурами. Все нейроны всех сетей постоянно взаимодействуют друг с другом. Это подтверждают многие исследователи.

Но все-таки можно выделить некоторые области в сером веществе мозга, которые более специализируются на отдельных задачах.
Зоны мозга нейрофизиологи выделяют следующие:
- Затылочная зона.
- Височная - отвечает за обоняние и вкус. Два эти чувства сильно взаимосвязаны.
- Зрительная зона. Тут расшифровываются сигналы, поступающие от глаз.
- Теменная - это так называемая зона кожно-мышечной чувствительности.
- Лобная доля - это сознательное поведение человека, его установки и трудовая деятельность. Задняя часть лобной доли - двигательный центр.
Функции больших полушарий мозга, как видим, распределены по зонам. Некоторые области имеют несколько функций. Например, руки связаны в больших полушариях с двумя зонами - двигательной и чувствительной.
И если при черепно-мозговой травме будет повреждена какая-либо из указанных областей, то функция этой зоны пострадает или совершенно пропадет. Восстановить утраченную функцию можно в том случае, если другая часть мозга - та, где находились нейроны, связанные с поврежденными тканями, сможет взять на себя всю работу утраченного центра.
Функции коры
Итак, каковы функции коры больших полушарий? Кора мозга отвечает за условные рефлексы, сформированные в процессе накопления опыта. Также в коре проходят все высшие психические процессы. Здесь сосредоточены зоны памяти, речи, мышления. Это более поздняя биологическая структура по сравнению с древним центральным мозгом, и она плохо изучена. Но известно, что наша личность и особенности характера, способность к усвоению и анализированию информации заложены именно в коре.
Большую роль играют в формировании навыков и привычек ассоциативные области. Можно сказать, утрируя информацию, что самая основная функция коры коры больших полушарий именно ассоциативная. Ведь на основе этих механизмов формируется и личность.
Ассоциативных областей 3:
- теменно-затылочно-височная;
- префронтальная ассоциативная;
- лимбическая.
Совместная работа этих центров обеспечивает всесторонний анализ поступающей извне информации. Без этих высших центров человек не смог бы целенаправленно выполнять работу.
Двигательная активность
Важнейшая функция больших полушарий - физическая активность. В передних отделах предцентральной извилины находится центр, где локализованы области проекции ступней и голеней. В средней части этой извилины находятся клетки, работающие с сигналами верхних конечностей, а самая глубокая часть предцентральной извилины отвечает за работу мышц лица.

Слаженная работа рецепторов проводящих нервных путей и этих мозговых центров обеспечивает нам ходьбу, работу руками и другую двигательную активность. Причем это все контролируется автоматически. Спортсмен ведь уже не думает, как согнуть ногу во время бега. Достаточно только дать сигнал старта сознательно.
Память и речь
В формировании памяти играют роль медиальная височная зона и гиппокамп. Однако они не являются тем местом, где накопленная информация хранится. Это скорее служебные зоны. Считается, что человек запоминает все, что видел или слышал когда-то. Основная проблема заключается в способности воспроизведения информации и ее перекодирования в слова.

Область речи - это граница височной и теменной зон. Причем у человека различают 2 зоны: отвечающий за речевое восприятие центр Вернике и за само произношение центр Брока.
Как лучше запомнить информацию?
Одна из функций больших полушарий, как мы теперь понимаем, - это запоминание и воспроизведение закодированной информации в словах. Если держать в мыслях и постоянно повторять одни и те же слова, то информация останется только в зоне речи и через несколько дней исчезнет.
Чтобы более глубоко запомнить информацию, необходимо применять образное мышление, ассоциируя каждое абстрактное понятие с яркими объектами.

В глубинной памяти у нас сохраняются только те аспекты реальности, которые связаны с яркими впечатлениями и сильными продолжительными эмоциями. А эмоции у нас "базируются" глубоко в белом веществе - в миндалевидном теле. Функции больших полушарий связаны с чисто сознательными намерениями запомнить.
Стрессы и депрессии ухудшают способность мозга запоминать что-либо. Начинать учить материал в беспокойном или раздражительном состоянии попросту бесполезно.
Вывод
Что можно сказать о функциях больших полушарий? Все центры мозга тесно взаимосвязаны. Говоря о конкретных областях, ученые подразумевают скопление нейронов, которые больше других взаимосвязанных сетей участвуют в том или ином психическом процессе.
Формирование памяти, способность говорить и думать словами - это самый сложный психический процесс. На это уходит большое количество энергии, и речью занято множество нервных клеток.
Кора больших полушарий связана непосредственно с сознательными процессами, а подкорка - с бессознательными, глубинными частями личности, которое Фрейд называл "Оно".
Кора больших полушарий головного мозга представляет собой наиболее молодое образование центральной нервной системы.Деятельность коры больших полушарий основана на принципе условного рефлекса, поэтому ее называют условно-рефлекторной. Она осуществляет быструю связь с внешней средой и приспособление организма к изменяющимся условиям внешней среды.
Глубокие борозды делят каждое полушарие большого мозга на лобную, височную, теменную, затылочную доли и островок . Островок расположен в глубине сильвиевой борозды и закрыт сверху частями лобной и теменной долей мозга.
Кора большого мозга делится на древнюю (архиокортекс ), старую (палеокортекс ) и новую (неокортекс). Древняя кора, наряду с другими функциями, имеет отношение к обонянию и обеспечению взаимодействия систем мозга. Старая кора включает поясную извилину, гиппокамп. У новой коры наибольшее развитие величины, дифференциации функций отмечается у человека. Толщина новой коры 3-4 мм. Общая площадь коры взрослого человека 1700-2000 см 2 , а число нейронов — 14 млрд (если их расположить в ряд, то образуется цепь протяженностью 1000 км) — постепенно истощается и к старости составляет 10 млрд (более 700 км). В составе коры имеются пирамидные, звездчатые и веретенообразные нейроны.
Пирамидные нейроны имеют разную величину, их дендриты несут большое количество шипиков: аксон пирамидного нейрона идет через белое вещество в другие зоны коры или структуры ЦНС.
Звездчатые нейроны имеют короткие, хорошо ветвящиеся дендриты и короткий аксон, обеспечивающий связи нейронов в пределах самой коры большого мозга.
Веретенообразные нейроны обеспечивают вертикальные или горизонтальные взаимосвязи нейронов разных слоев коры.
Строение коры больших полушарий
В коре содержится большое количество глиальных клеток, выполняющих опорную, обменную, секреторную, трофическую функции.
Наружная поверхность коры разделена на четыре доли: лобную, теменную, затылочную и височную. Каждая доля имеет свои проекционные и ассоциативные области.
Кора большого мозга имеет шестислойное строение (рис. 1-1):
- молекулярный слой (1) светлый, состоит из нервных волокон и имеет небольшое количество нервных клеток;
- наружный зернистый слой (2) состоит из звездчатых клеток, определяющих длительность циркулирования возбуждения в коре головного мозга, т.е. имеющих отношение к памяти;
- слой пирамидных меток (3) формируется из пирамидных клеток малой величины и вместе со слоем 2 обеспечивает корко-корко- вые связи различных извилин мозга;
- внутренний зернистый слой (4) состоит из звездчатых клеток, здесь заканчиваются специфические таламокортикальные пути, т.е. пути, начинающиеся от рецепторов-анализаторов.
- внутренний пирамидный слой (5) состоит из гигантских пирамидных клеток, которые являются выходными нейронами, аксоны их идут в ствол мозга и спинной мозг;
- слой полиморфных клеток (6) состоит из неоднородных по величине клеток треугольной и веретенообразной формы, которые образуют кортикоталамические пути.
I — афферентные пути из таламуса: СТА — специфические таламические афференты; НТА — неспецифические таламические афференты; ЭМВ — эфферентные моторные волокна. Цифрами обозначены слои коры; II — пирамидный нейрон и распределение окончаний на нем: А — неспецифические афферентные волокна из ретикулярной формации и ; Б — возвратные коллатерали от аксонов пирамидных нейронов; В — комиссуральные волокна из зеркальных клеток противоположного полушария; Г — специфические афферентные волокна из сенсорных ядер таламуса
Рис. 1-1. Связи коры больших полушарий.
Клеточный состав коры по разнообразию морфологии, функций, формам связи не имеет себе равных в других отделах ЦНС. Нейронный состав, распределение по слоям в разных областях коры различны. Это позволило выделить в мозге человека 53 цитоархитектонических поля. Разделение коры большого мозга на цитоархитектонические поля более четко формируется по мере совершенствования ее функции в филогенезе.
Функциональной единицей коры является вертикальная колонка диаметром около 500 мкм. Колонка - зона распределения разветвлений одного восходящего (афферентного) таламокортикального волокна. Каждая колонка содержит до 1000 нейронных ансамблей. Возбуждение одной колонки тормозит соседние колонки.

Восходящий путь проходит через все корковые слои (специфический путь). Неспецифический путь также проходит через все корковые слои. Белое вещество полушарий расположено между корой и базальными ганглиями. Оно состоит из большого количества волокон, идущих в разных направлениях. Это проводящие пути конечного мозга. Различают три вида путей.
- проекционный — связывает кору с промежуточным мозгом и другими отделами ЦНС. Это восходящие и нисходящие пути;
- комиссуральный - его волокна входят в состав мозговых комиссур, которые соединяют соответствующие участки левого и правого полушарий. Входят в состав мозолистого тела;
- ассоциативный - связывает участки коры одного и того же полушария.
Зоны коры больших полушарий
По особенностям клеточного состава поверхность коры подразделяют на структурные единицы следующего порядка: зоны, области, подобласти, поля.
Зоны коры головного мозга разделяются на первичные, вторичные и третичные проекционные зоны. В них расположены специализированные нервные клетки, к которым поступают импульсы от определенных рецепторов (слуховых, зрительных и т.д.). Вторичные зоны представляют собой периферические отделы ядер анализаторов. Третичные зоны получают обработанную информацию от первичных и вторичных зон коры больших полушарий и играют важную роль в регуляции условных рефлексов.
В сером веществе коры больших полушарий различают сенсорные, моторные и ассоциативные зоны:
- сенсорные зоны коры больших полушарий -
участки коры, в которых располагаются центральные отделы анализаторов:
зрительная зона — затылочная доля коры больших полушарий;
слуховая зона — височная доля коры больших полушарий;
зона вкусовых ощущений — теменная доля коры больших полушарий;
зона обонятельных ощущений — гиппокамп и височная доля коры больших полушарий.
Соматосенсорная зона находится в задней центральной извилине, сюда приходят нервные импульсы от проприорецепторов мышц, сухожилий, суставов и импульсы от температурных, тактильных и других рецепторов кожи;
- моторные зоны коры больших полушарии - участки коры, при раздражении которых появляются двигательные реакции. Располагаются в передней центральной извилине. При ее поражении наблюдаются значительные нарушения движения. Пути, по которым импульсы идут от больших полушарий к мышцам, образуют перекрест, поэтому при раздражении моторной зоны правой стороны коры возникает сокращение мышц левой стороны тела;
- ассоциативные зоны - отделы коры, находящиеся рядом с сенсорными зонами. Нервные импульсы, поступающие в сенсорные зоны, приводят к возбуждению ассоциативных зон. Особенностью их является то, что возбуждение может возникать при поступлении импульсов от различных рецепторов. Разрушение ассоциативных зон приводит к серьезным нарушениям обучения и памяти.
Речевая функция связана с сенсорными и двигательными зонами. Двигательный центр речи (центр Брока) находится в нижней части левой лобной доли, при его разрушении нарушается речевая артикуляция; при этом больной понимает речь, но сам говорить не может.
Слуховой центр речи (центр Вернике) расположен в левой височной доле коры больших полушарий, при его разрушении наступает словесная глухота: больной может говорить, излагать устно свои мысли, но не понимает чужой речи; слух сохранен, но больной не узнает слов, нарушается письменная речь.
Речевые функции, связанные с письменной речью — чтение, письмо, — регулируются зрительным центром речи, расположенным на границе теменной, височной и затылочной долей коры головного мозга. Его поражение приводит к невозможности чтения и письма.
В височной доле находится центр, отвечающий за запоминание слое. Больной с поражением этого участка не помнит названия предметов, ему необходимо подсказывать нужные слова. Забыв название предмета, больной помнит его назначение, свойства, поэтому долго описывает их качества, рассказывает, что делают с этим предметом, но назвать его не может. Например, вместо слова «галстук» больной говорит: «это то, что надевают на шею и завязывают специальным узлом, чтобы было красиво, когда идут в гости».
Функции лобной доли:
- управление врожденными поведенческими реакциями при помощи накопленного опыта;
- согласование внешних и внутренних мотиваций поведения;
- разработка стратегии поведения и программы действия;
- мыслительные особенности личности.
Состав коры больших полушарий
Кора больших полушарий головного мозга является высшей структурой ЦНС и состоит из нервных клеток, их отростков и нейроглии. В составе коры имеются звездчатые, веретенообразные и пирамидные нейроны. Благодаря наличию складок кора имеет большую поверхность. Выделяют древнюю кору (архикортекс) и новую кору (неокортекс). Кора состоит из шести слоев (рис. 2).

Рис. 2. Кора больших полушарий головного мозга
Верхний молекулярный слой образован в основном дендритами пирамидных клеток нижележащих слоев и аксонами неспецифических ядер таламуса. На этих дендритах формируют синапсы афферентные волокна, приходящие от ассоциативных и неспецифических ядер таламуса.
Наружный гранулярный слой образован мелкими звездчатыми клетками и частично малыми пирамидными клетками. Волокна клеток этого слоя расположены преимущественно вдоль поверхности коры, формируя кортикокортикальные связи.
Слой пирамидных клеток малой величины.
Внутренний гранулярный слой, образованный звездчатыми клетками. В нем заканчиваются афферентные таламокортикальные волокна, начинающиеся от рецепторов анализаторов.
Внутренний пирамидный слой состоит из крупных пирамидных клеток, участвующих в регуляции сложных форм движения.
Мультиформный слой состоит из верстеновидных клеток, образующих кортикоталамические пути.
По функциональной значимости нейроны коры подразделяют на сенсорные , воспринимающие афферентные импульсы от ядер таламуса и рецепторов сенсорных систем; моторные , посылающие импульсы к подкорковым ядрам, промежуточному, среднему, продолговатому мозгу, мозжечку, ретикулярной формации и спинному мозгу; и промежуточные , осуществляющие связь между нейронами коры больших полушарий. Нейроны коры больших полушарий находятся в состоянии постоянного возбуждения, не исчезающего и во время сна.
В кору больших полушарий, к сенсорным нейронам поступают импульсы от всех рецепторов организма через ядра таламуса. И каждый орган имеет свою проекцию или корковое представительство, расположенное в определенных областях больших полушарий.
В коре больших полушарий имеется четыре чувствительные и четыре двигательные области.
Нейроны двигательной коры получают афферентную импульсацию через таламус от мышечных, суставных и кожных рецепторов. Основные эфферентные связи двигательной коры осуществляются через пирамидные и экстрапирамидные пути.
У животных наиболее развита лобная область коры и ее нейроны участвуют в обеспечении целенаправленного поведения. Если удалить эту долю коры, животное становится вялым, сонливым. В височной области локализуется участок слуховой рецепции, и сюда поступают нервные импульсы от рецепторов улитки внутреннего уха. Область зрительной рецепции находится в затылочных долях коры головного мозга.
Теменная область, внеядерная зона, играет важную роль в организации сложных форм высшей нервной деятельности. Здесь расположены рассеянные элементы зрительного и кожного анализаторов, осуществляется межанализаторный синтез.
Рядом с проекционными зонами располагаются ассоциативные зоны, которые осуществляют связь между сенсорной и двигательной зонами. Ассоциативная кора принимает участие в конвергенции различных сенсорных возбуждений, позволяющей осуществлять сложную обработку информации о внешней и внутренней среде.